|
Corresponding author: Patrick Okechukwu ( patrickn@ucsiuniversity.edu.my ) Academic editor: Georgi Momekov
© 2020 Patrick Okechukwu, Mridula Sharma, Wen Hui Tan, Hor Kuan Chan, Kavita Chirara, Anand Gaurav, Mayasah Al-Nema.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Okechukwu P, Sharma M, Tan WH, Chan HK, Chirara K, Gaurav A, Al-Nema M (2020) In-vitro antidiabetic activity and in-silico studies of the binding energies of palmatine and the standard compounds with the three receptors of alpha amylase, alpha glucosidase, and DPP-IV enzyme. Pharmacia 67(4): 363-371. https://doi.org/10.3897/pharmacia.67.e58392
|
Abstract
Palmatine a protoberberine alkaloid has been previously reported to possess in vivo antidiabetic and antioxidant property. The aim of the experiment is to evaluate the in vitro antidiabetic activity and in-silico studies of the binding energies of Palmatine, acarbose, and Sitagliptin with the three enzymes of alpha-amylase, alpha-glucosidase, and dipeptidyl peptidase-IV (DPP-IV). The in vitro antidiabetic study was done by evaluating the inhibitory effect of palmatine on the activities of alpha-amylase, alpha-glucosidase, and DPP-IV. Acarbose, and sitagliptin was used as standard drug. The molecular docking study was performed to study the binding interactions of palmatine with alpha-glucosidase, a-amylase, and DPP-IV. The binding interactions were compared with the standard compounds Sitagliptin and acarbose. Palmatine with IC50 (1.31 ± 0.27 µM) showed significant difference of (< 0.0001) higher inhibiting effect on alpha-amylase and weak inhibiting effect on alpha-glucosidase enzyme with IC50 (9.39 ± 0.27 µM) and DPP-IV with IC50 (8.7 ± 1.82 µM). Palmatine possess inhibition effect on the three enzymes.
Keywords
Alpha-glucosidase, alpha-amylase, dipeptidyl peptidase-IV, diabetes mellitus, palmatine
Introduction
Diabetes mellitus (DM) is a metabolic multiple etiology disease associated with high blood sugar levels (
Palmatine, is a bioactive protoberberine alkaloid, which was isolated from Coscinium fenestratum (CF) stem extract. CF belongs to the plant family name Menispermaceae and commonly known as ‘tree turmeric’. Previous report has shown that the stem extract of CF and Palmatine possess in vivo antidiabetic and antioxidant properties. This study was aimed to evaluate the in-vitro antidiabetic activity and in-silico studies of the binding energies of palmatine and the standard compounds with the three active centers of alpha-amylase, alpha-glucosidase, and DPP-IV enzymes.
Materials
Abbreviation’s
ADT = AutoDockTools, DM = Diabetes melltius; DNS = 3,5-Dintirosalicylic acid; DPP-IV = Dipeptidyl peptidase-IV; GIP = Glucose-dependent insulin tropic polypeptide; GLP-1 = glucagon-like peptide 1; NaOH = sodium hydroxide.
Chemicals
Palmatine (Sigma Aldrich, USA), alpha-glucosidase from Saccharomyces cerevisiae (E.C. 3.2.1.20) (Sigma Aldrich, USA), alpha-amylase from porcine amylase ((E.C. 3.2.1.1) (Sigma-Aldrich, USA) DPP-IV enzyme (Sinar Scientific, Malaysia), starch (Friendmann Schmdit, Austria), sucrose (EMD, Millipore Chemicals, France), 6N HCL (Sinar Scientific, Malaysia), Sodium phosphate buffer (Sigma Aldrich, USA), 3,5-dinitrosalicylic acid (Systrem, Malaysia), sodium potassium tartrate tetrahydrate (Hopkins and Williams, UK), Sodium hydroxide (Systrem, Malaysia), Rebaudioside (Sigma Aldrich, USA), Tris buffer (Bio-Rad, United States), 25% glacial acetic acid (Merck, Germany), Acarbose (Sigma-Aldrich, USA), Sitagliptin (Januvia, MSD Pharmaceutical Pvt. Ltd., India), Glimepiride (Amaryl, Sanofi India Pvt. Ltd., India), Metformin (Glucophage XR, Merck, Malaysia).
Equipment
Vortex mixer (LMS CO., LTD, Japan); Centrifuge machine (Zentrifugen, Germany); pH meter (Mettler Tolledo, Switzerland); UV-Vis spectrophotometer (SECOMAM, France); Water bath (Copens Scientific (M) Sdn Bhd, Malaysia); Electronic balance (Mettler Toledo, Switzerland); FLUOstar Omega Plate Reader (Thermofisher, USA).
Methodology
In-vitro Anti-diabetic studies
Alpha-Amylase inhibiting activity
The inhibiting activity of alpha-amylase was performed according to the method by
Alpha-Glucosidase inhibiting activity
The alpha-glucosidase inhibiting activity was performed according to a method described by
Dipeptidyl peptidase IV inhibiting activity (DPP-IV)
The inhibiting activity of dipeptidyl peptidase IV (DPP-IV) was performed according to the method by
In-silico studies
The molecular docking study was performed using AutoDock Vina 1.1.2. The X-ray crystallographic structures of the target receptors, alpha – glucosidase (PDB ID: 3W37), alpha – amylase (PDB ID: 4W93) and DPP-IV (PDB ID: 4A5S), bound with their co-crystallized ligands were retrieved from the protein data bank (PDB) at resolutions of 1.70 Å, 1.35 Å and 1.62 Å, respectively. The protein structures were prepared for docking using Biovia Discovery Studio (DS) and AutoDock Tools (ADT) 1.5.6. All the water molecules, co-crystallized ligands and other heteroatoms were removed and polar hydrogen atoms, as well as Gasteiger charges, were added. Following the protein preparation, the structures of palmatine and the standard compounds, acarbose and Sitagliptin, were downloaded from PubChem and converted into their respective 3D structures using Biovia DS. The ligands were imported into AutoDock where ADT was used to add charges, set the rotatable bonds, and allow all the torsions to rotate for the ligands. Then, the amino acid residues of the prepared protein structures that involved in the interactions with the co-crystallized ligands were determined in which the binding site of each protein structure was identified based on the determined residues. ADT was used to determine the Grid Box that covers the entire binding site of the protein. The coordinates and size of the Grid Box were saved in the input parameter file. Finally, the prepared ligands were docked into the active sites of the prepared proteins’ structures. All the docking parameters have remained as default settings. The binding energies of the ligands were recorded and compared (the more negative value indicates the higher affinity). The binding poses of the ligands and the mode of interactions of the protein-ligand complexes were studied thoroughly using Biovia DS.
Statistical analysis
All data were presented as mean (±) standard deviation (SD) in triplicates (n = 6) using the XL. STAT 7.0. The data were statistically analyzed by One-Way ANOVA followed by Dunnet’s Test.Values were considered statistically significant and denoted as; p < 0.05 = a; p < 0.01 = b; p < 0.001 = c; p < 0.0001 = d and data which were not found significantly different from each other, denoted by same superscript letter.
Results
In-vitro anti-diabetic activities
Alpha-amylase enzyme inhibiting activity
The effect of palamtine on the alpha-amylase activity was investigated using starch substrate. Acarbose was used as psitive control and glimepiride and metformin was used as standard drug. On the basis of IC50, Palmatine with IC50 (1.31 ± 0.27 µM) showed significant (< 0.0001) higher inhibiting effect on alpha-amylase followed by metformin (2.5 ± 0.47 µM), acarbose (2.94 ± 0.36 µM) and glimepiride (4.98 ± 0. 47 µM) as shown Figure

Alpha-amylase enzyme inhibting activity (IC50(µM) of palmatine, glimepiride, metformin and standard drug(acarbose). The result are shown as Mean ± standard deviation in triplicates (n = 3). The data were statistically analyzed by One-way ANOVA followed by Dunnet’s post hoc test. Graph showed that acarbose(a) vs. glimepiride = p < 0.001 = c; acarbose vs. metformin = p < 0.01 = b; acarbose vs. palmatine = p < 0.0001 = d. Glimepiride vs. Metformin = p < 0.001 = c, glimepiride vs. palmatine = p < 0.0001 = d, metformin vs. palmatine = p < 0.0001 = d.
Alpha-glucosidase enzyme inhibiting activity
The effect of palamtine on the alpha-glucosidase enzyme activity was investigated using sucrose as substrate. Acarbose was used as positive control drug, glimepiride and metformin was used as standard drugs. On the basis of IC50, palmatine had a weak inhibiting effect on alpha-glucosidase enzyme with IC50 (9.39±0.27µM). Metformin and glimepiride had similar inhibtiory effect with IC50 (9.5 ± 1.11 µM and 9.9 ± 0.22 μM). However, acarbose showed a very strong inhibtiory effect with IC50 (1.31 ± 0.2 µM) as shown in Figure

Alpha-glucosidase enzyme inhibting activity ((IC50(µM) of palmatine, glimepiride, metformin and standard drug(acarbose). The result are shown as Mean ± standard deviation in triplicates (n = 3). The data were statistically analyzed by One-way ANOVA followed by Dunnet’s post hoc test. Graph showed that acarbose(a) vs. glimepiride = p < 0.0001 = d; acarbose vs. metformin = p < 0.0001 = d; acarbose vs. palmatine = p < 0.0001 = d. Glimepiride vs. metformin = No significant = d, glimepiride vs. palmatine = No significant = d and metformin vs. palmatine = No significant = d.
Dippetidyl peptidase enzyme (DPP IV) Inhibiting activity
The effect of palamtine on the dippetidyl peptidase enzyme activity was investigated using rebaudioside as a substrate. Sitagliptin was used as positive drug, glimepiride and metformin was used as standard drug. On the basis of IC50, palmatine had a weak inhibiting effect on DPP-IV with IC50 (8.7 ± 1.82 µM) and sitagliptin showed a very strong inhibtiory effect with IC50 (1.07 ± 0.23 µM) as shown in Figure

DPP-IV enzyme inhibting activity ((IC50(µM) of glimepiride, metformin, palmatine and standard drug(sitagliptin). The result are shown as Mean ± standard deviation in triplicates (n = 3). The data were statistically analyzed by One-way ANOVA followed by Dunnet’s post hoc test. Graph showed that sitagliptin (a) vs. palmatine = p < 0.0001 = d. Glimepiride and metformin = NA = No activity.
In-silico studies
The docking study of palmatine and the standard compounds, acarbose and sitagliptin, allowed to display the affinity and the best binding pose of the respective compounds within the active sites of alpha-glucosidase, alpha-amylase and DPP-IV in addition to the elucidation of the interactions and the amino acids involved in the binding. The results of docking studies are presented in Table
The binding energies of palmatine and the standard compounds with the three active centers of alpha-amylase, alpha-glucosidase and DPP-IV enzymes (Table shows the result of the binding energies of palmatine and the standard drug acarbose and sitagliptin).
| Compound | Structure | Binding Energy (Kcal/mol) | ||
|---|---|---|---|---|
| alpha-glucosidase | alpha-amylase | DPP-IV | ||
| Palmatine |
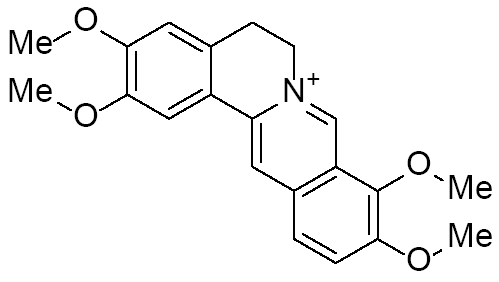
|
-6.4 | -7.8 | -8.0 |
| Acarbose |
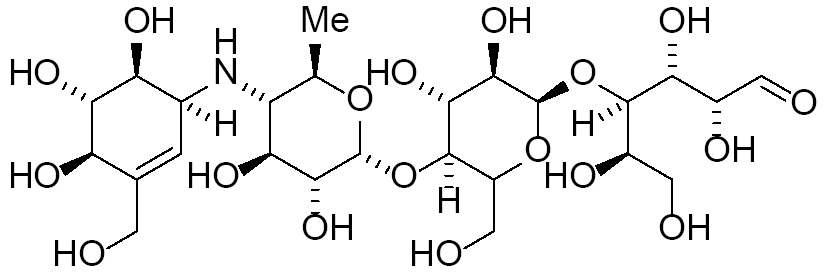
|
-8.3 | -8.8 | -9.4 |
| Sitagliptin |
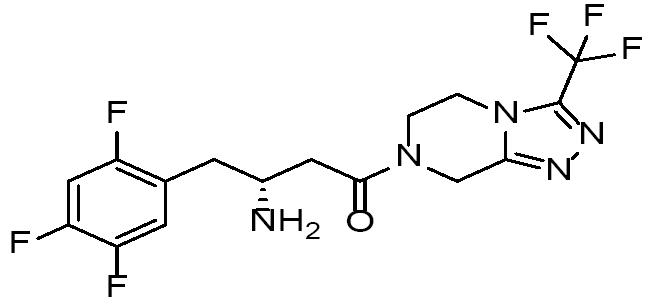
|
-8.7 | -8.7 | -9.2 |
Discussion
Hyperglycemia is categorized as a common metabolic abnormality in diabetes mellitus. Hyperglycemia is a condition in which an excessive quantity of glucose circulates in the blood plasma due to insufficient secretion or impaired action of insulin (
According to the docking results, acarbose has shown the highest binding affinity (binding energy -8.8 kcal/mol) to the alpha-amylase. The high affinity is attributed to the hydrogen bond interactions between the ligand and the active site residues of the receptor. As can be seen in Figure

In contrast to acarbose, no hydrogen bond interactions were observed between palmatine and alpha-amylase. The isoquinoline group of palmatine displays π-π staked interactions with the aromatic ring of TRP59. Additionally, the tetrahydronaphthalene group displays another π-π staked interaction with the aromatic residue TYR62 (Figure

The binding mode of acarbose within the active site of alpha-glucosidase receptor showed that the ligand forms two hydrogen bond interactions with the receptor. These hydrogen bonds form between the hydroxyl groups of the ligand and the residue MET470 and SER497, respectively, in which the amino acids residues act as hydrogen bond donors. Moreover, the methyl group of the ligand interacts with the aromatic rings of PHE236 and PHE601 (Figure


Both acarbose and palmatine displayed similar interactions with alpha-glucosidase. However, acarbose exhibited a higher affinity to the receptor (binding energy -8.3 Kcal/mol) than palmatine (binding energy -6.4 Kcal/mol). The higher affinity can be attributed, to the hydrogen bond interactions with two different residues MET470 and SER497 which were observed with acarbose in contrast, to the hydrogen bond interactions with only one residue SER497 which, were observed with palmatine.
Sitagliptin was used as a standard compound where it exhibited high affinity towards DPP-IV receptor (binding energy -9.2 Kcal/mol). The ligand involved in several interactions with DPP-IV in which the two fluorine groups of the phenyl ring and the carbonyl group form four hydrogen bonds with the residues ARG358 and ARG125, respectively. The ligand acts as hydrogen bond acceptor where it accepted the hydrogen atoms from ARG. Additionally, the phenyl ring of the ligand displays π-π staked interactions with the aromatic residue PHE357 and the methyl group interacts with the aromatic residues TRP659, TYR662 and TYR666 (Figure


The binding interactions of sitagliptin (four hydrogen bonds and the aromatic interactions) contribute to the strength of binding and the high affinity of the ligand towards the DPP-IV receptor in comparison to palmatine which involved in only one hydrogen bond and three aromatic interactions with the same residue of the receptor.
Conclusion
The results of the present study prove that palmatine is effective alpha-amylase, alpha-glucosidase and DPP-IV inhibitors, which may be helpful to reduce the postprandial glucose level. The binding interactions of palmatine with the alpha-amylase, alpha-glucosidase and DPP-IV identified through docking studies helped to shed light on the mechanism of its binding with these three antidiabetic targets. These finding can be extremely useful in designing new semisynthetic derivatives of palmatine as antidiabetic compounds.
Acknowledgement
The authors acknowledge UCSI University for providing the fund and facility for this research work.
References
- Ahmed F, Chandra J, Timmaiah N (2009) An In vitro study on the inhibitory activities of Eugenia jambolana seeds against carbohydrate hydrolyzing enzymes. Journal of Young Pharmacists 1(4): 1–317. https://doi.org/10.4103/0975-1483.59320
- Akuba B, Ozougwu V (2018) In vitro Inhibition of Carbohydrate Metabolizing Enzymes and in vivo Anti-hyperglycaemic Potential of Methanol Extract of Desmodium velutinum Leaves. Research Journal of Medicinal Plants 12(1): 48–56. https://doi.org/10.3923/rjmp.2018.48.56
- Beidokhti M, Lobbens E, Rasoavaivo P, Staerk D, Jäger A (2018) Investigation of medicinal plants from Madagascar against DPP-IV linked to type 2 diabetes. South African Journal of Botany 115: 113–119. https://doi.org/10.1016/j.sajb.2018.01.018
- Cho N, Shaw J, Karuranga S, Huang Y, da Rocha Fernandes J, Ohlrogge A, Malanda B (2018) IDF Diabetes Atlas: Global estimates of diabetes prevalence for 2017 and projections for 2045. Diabetes Research and Clinical Practice 138: 271–281. https://doi.org/10.1016/j.diabres.2018.02.023
- González-Montoya M, Hernández-Ledesma B, Mora-Escobedo R, Martínez-Villaluenga C (2018) Bioactive Peptides from Germinated Soybean with Anti-Diabetic Potential by Inhibition of Dipeptidyl Peptidase-IV, α-Amylase, and α-Glucosidase Enzymes. International journal of molecular sciences 19(10): e2883. https://doi.org/10.3390/ijms19102883
- Kharroubi AT, Darwish HM (2015) Diabetes mellitus: The epidemic of the century. World journal of diabetes 6(6): 850–867. https://doi.org/10.4239/wjd.v6.i6.850
- Kidane Y, Bokrezion T, Mebrahtu J, Mehari M, Gebreab Y, Fessehaye N, Achila O (2018) In Vitro Inhibition of α-Amylase and α-Glucosidase by Extracts from Psiadia punctulata and Meriandra bengalensis. Evidence-Based Complementary and Alternative Medicine 1–9. https://doi.org/10.1155/2018/2164345
- Kim B, Kim H, Choi I, Kim J, Jin C, Han A (2018) DPP-IV Inhibitory Potentials of Flavonol Glycosides Isolated from the Seeds of Lens culinaris: In Vitro and Molecular Docking Analyses. Molecules 23(8): e1998. https://doi.org/10.3390/molecules23081998
- Kortykowski M, McDonnell ME, Umpierrez GE, Zonszein J (2012) Patient Guide to Managing Hyperglycemia (High Blood Sugar) in the Hospital. The Journal of Clinical Endocrinology Metabolism 97(1): 27A–28A. https://doi.org/10.1210/jcem.97.1.zeg27a
- Lin SR, Chang CH, Tsai MJ, Cheng H, Chen JC, Leong MK, Weng CF (2019) The perceptions of natural compounds against dipeptidyl peptidase 4 in diabetes: from in silico to in vivo. Therapeutic advances in chronic disease 10: e2040622319875305. https://doi.org/10.1177/2040622319875305
- McDougall G, Shpiro F, Dobson P, Smith P, Blake A, Stewart D (2005) Different Polyphenolic Components of Soft Fruits Inhibit α-Amylase and α-Glucosidase. Journal of Agricultural and Food Chemistry 53(7): 2760–2766. https://doi.org/10.1021/jf0489926
- Meneses M, Silva B, Sousa M, Sá R, Oliveira P, Alves M (2015) Antidiabetic Drugs: Mechanisms of Action and Potential Outcomes on Cellular Metabolism. Current Pharmaceutical Design 21(25): 3606–3620. https://doi.org/10.2174/1381612821666150710145753
- Pandithurai M, Murugesan S, Bhuvaneswari S, Thennarasan S (2015) In-vitro alpha-amylase and alpha glucosidase inhibition activity of methanolic extract of marine brown alga Spatoglossum asperum. Advances in Pharmaceutics 4(5).
- Rangel É, Rodrigues C, de Sá J (2019) Micro- and Macrovascular Complications in Diabetes Mellitus: Preclinical and Clinical Studies. Journal of Diabetes Research 2019(2161085): 1–5. https://doi.org/10.1155/2019/2161085
- Salehi M, Aulinger B, D’Alessio D (2012) Effect of Glycemia on Plasma Incretins and the Incretin Effect During Oral Glucose Tolerance Test. Diabetes 61(11): 2728–2733. https://doi.org/10.2337/db11-1825
- Somtimuang C, Olatunji O, Ovatlarnporn C (2018) Evaluation of in Vitro α-Amylase and α-Glucosidase Inhibitory Potentials of 14 Medicinal Plants Constituted in Thai Folk Antidiabetic Formularies. Chemistry & Biodiversity 15(4): e1800025. https://doi.org/10.1002/cbdv.201800025
- Tay Y, Bakar M, Azmi M, Saad N, Awang K, Litaudon M, Kassim M (2020) Inhibition of Carbohydrate Hydrolysing Enzymes, Antioxidant Activity and Polyphenolic Content of Beilschmiedia Species Extracts. IOP Conference Series: Materials Science and Engineering 716: 012007. https://doi.org/10.1088/1757-899X/716/1/012007
- Telagari M, Hullatti K (2015) In-vitro α-amylase and α-glucosidase inhibitory activity of Adiantum caudatum Linn. and Celosia argentea Linn. extracts and fractions. Indian journal of pharmacology 47(4): 425–429. https://doi.org/10.4103/0253-7613.161270
- Thakare V, Shende S, Shirure P, Swami O (2017) Role of conventional oral antidiabetic drugs in management of type 2 diabetes mellitus. International Journal of Research in Medical Sciences 5(3): e749. https://doi.org/10.18203/2320-6012.ijrms20170619
- Unuofin J, Otunola G, Afolayan A (2018) In vitro α-amylase, α-glucosidase, lipase inhibitory and cytotoxic activities of tuber extracts of Kedrostis africana (L.) Cogn. Heliyon 4(9): e00810. https://doi.org/10.1016/j.heliyon.2018.e00810
- Yin Z, Zhang W, Feng F, Zhang Y, Kang W (2014) α-Glucosidase inhibitors isolated from medicinal plants. Food Science and Human Wellness 3(3–4): 136–174. https://doi.org/10.1016/j.fshw.2014.11.003