Research Article |
|
Corresponding author: Yara Al Tall ( yraltall@just.edu.jo ) Academic editor: Rumiana Simeonova
© 2023 Yara Al Tall, Baker Al-Nassar , Ahmad Abualhaijaa , Salsabeel H. Sabi, Ammar Almaaytah.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Al Tall Y, Al-Nassar B, Abualhaijaa A, Sabi SH, Almaaytah A (2023) The design and functional characterization of a novel hybrid antimicrobial peptide from Esculentin-1a and melittin. Pharmacia 70(1): 161-170. https://doi.org/10.3897/pharmacia.70.e97116
|
Abstract
Antimicrobial agents are one of the most widely used drugs in medicine. In the last fifty years, the misuse of these agents caused the emergence of resistant strains of bacteria that led to an increase in life-threatening infections. The need to develop new agents has become a priority, and antimicrobial peptides attained high consideration. The antimicrobial activities of a novel In-house designed hybrid cationic peptide (BKR1) were studied against different strains of Gram-negative bacteria. This was done using the broth dilution method as outlined by the Clinical and Laboratory Institute (CLSI). Checkerboard assy was employed to investigate the synergistic activity of BKR1 peptide with four antibiotics (Levofloxacin, chloramphenicol, rifampicin, and ampicillin). Finally, the cytotoxicity of BKR1 was evaluated against human blood cells and mammalian kidney cells (Vero cells). BKR1 displayed bactericidal activity against tested strains of Gram-negative bacteria, with zero hemolytic effects. It also acts as a strong adjuvant with levofloxacin, chloramphenicol, and rifampicin against resistant strains of P. aeruginosa and E. coli. This study represents the design and elucidation of the antimicrobial activities of a novel hybrid antimicrobial peptide named (BKR1). Our results indicate thar BKR1 is a promising candidate to treat resistant infectious diseases individually or as an adjuvant with conventional antibiotics.
Keywords
Antimicrobial resistance, hybrid peptide, antibiotic adjuvant, bactericidal activity, hemolytic activity
Introduction
Infectious diseases are one of the leading causes of mortality all around the world (Waldman et al. 2016). Although many antimicrobial agents have been developed in previous decades, the misuse of these treatments led to severe resistance among several microorganisms, which are now called multi-drug resistant bacteria (MDRB) (
The combination antimicrobial therapy approach has gained interest to treat resistant bacterial infections, including antibiotic-antibiotic or antibiotic-adjuvant combinations (
Cationic antimicrobial peptides (AMPs) are considered potential antimicrobial agents and/or adjuvants to antibiotics (
AMPs are generally short, less than 50 amino acids in length (
In this study, we have designed a novel hybrid peptide based on the amino acid sequence of both Esculentin-1a and melittin. Esculentin-1a exhibits strong antimicrobial activity and was found in the skin of many amphibians as a type of an innate defense mechanism (
The newly designed hybrid peptide named BKR1 carries a higher positive charge when compared with the parent peptides; we deduced that this modification would enhance BKR1’s antimicrobial activity against bacterial strains (
Materials and methods
Materials, chemicals and bacterial strains
All strains used in this study were obtained from American Type Tissue Culture Collection (ATCC; Manassas, VA, USA). The four strains used in this study are Pseudomonas aeruginosa (ATCC 27853 and BAA-2114), Escherichia coli (ATCC 25992 and BAA-2452). All chemicals were purchased from Sigma Aldrich (St. Louis, MO, USA) unless stated otherwise.
Peptide design, molecular modelling and in silico analysis
Rational design of the new hybrid peptide was performed using online bioinformatics software to achieve the optimal range in physiochemical properties. The Hierarchical Neural Network software (HNN) from the Network Protein Sequence Analysis (NPS) server was used to predict the secondary structure and calculate the helicity percentages of the parent peptides and the hybrid peptide. The physicochemical properties were predicted using ProtParam/ExPASy server (
Peptide design, synthesis, and purification
BKR1, a hybrid peptide of 21 amino acids (NH2-VSKLAKKIKNLKNVKKSWKRQ-COOH) was synthesized following standard Fmoc solid-phase protocols on Wang resin. Peptide elongation was performed using standard HBTU coupling chemistry in dimethylformamide (DMF) solvent with a fourfold molar excess of diisopropyl ethylamine (DIEA) in N-methyl-2-pyrrolidone (NMP) and a threefold molar excess of each Fmoc-protected amino acid or 2-(6-methoxynaphthalen-2-yl) propanoic acid. BKR! was cleaved from the resin, using 95% trifluoroacetic acid (TFA), 2.5% triisopropylsilane, and 2.5% thioanisole (3 h, room temperature), and precipitated using cold (–20 °C) diethyl ether. Reverse phase high-performance liquid chromatography (RP-HPLC) was used for purification of BKR1 using a C18 internsil ODS-SP column, the column was eluted with acetonitrile / H2O-TFA gradient at flow rate of 1.0 ml/minute. The identification of BKR! was confirmed by mass analysis and through the employment of electrospray ionization mass spectrometry (ESI-MS).
Bacterial susceptibility assay
Antimicrobial activities of the peptide alone, the antibiotics alone and in combination were tested using the broth dilution method as described by the Clinical and Laboratory Standards Institute guidelines and as performed previously (
Briefly, four bacterial strains including a control and resistant clinical isolate of P. aeruginosa (ATCC 27853 and ATCC BAA-2114) in addition to a control and resistant clinical isolate of E. coli (ATCC 25922 and ATCC BAA-2452) were grown in Muller Hinton broth (MHB) medium. The initial concentration of the bacterial culture (1.5 × 108 CFU/mL) were adjusted using 0.5 McFarland turbidity standard, and spectrophotometry at λ = 600 nm. The used concentration was diluted 100-fold to reach a value of 106 CFU/mL. The bacterial suspension of the diluted concentration was distributed over 96-well plates. Each plate had a range different concentration of one of the four tested antibiotics (LVX, CHL, RIF, and AMP) that were determined according to previous analysis of the MIC values of each antibiotic against the target strain ranging from (1–100 µM), or different concentrations of the hybrid peptide. The growth control consisted of bacterial suspension without any concentration of an antimicrobial agent, while the negative control consisted of MHB broth alone.
The plates were incubated overnight, and the bacterial growth was evaluated by measuring the absorbance at λ = 600 nm using an Enzyme-Linked Immunosorbent Assay (ELISA) microplate reader (EpochTM; BioTek, Winooski, VT, USA). The minimum inhibitory concentration (MIC) values of the antimicrobial agent (antibiotics or hybrid peptide) were assessed by the bacterial growth at this stage since the MIC value is defined as The minimum inhibitory concentration (MIC) is the lowest antimicrobial concentration at which no visible microbial growth can be detected with the naked eye or where there is no difference in absorbance between the negative control and the test concentration (
Additonally, the minimum bactericidal concentration (MBC) which is defined as “the lowest concentration of antimicrobial that will prevent the growth of an organism after subculture on to antibiotic free media” (Mohseni et al. 2014), was determined by taking samples of 10 µL from turbidity-free wells, and spreading them over agar plates, followed by incubation for 24–48 h. Colonies were then counted and compared with growth controls.
The MIC and the MBC values for the four antibiotics and the hybrid peptide were tested alone and in combination against different bacterial strains as mentioned above. All experiments were made in triplicates. The checkerboard assay was adopted to check the synergistic activities between the hybrid peptide and the antibiotics.
Synergistic checkerboard assay
The checkerboard assay employs different concentrations of antimicrobial agents to check their synergistic activities against a specific bacterial strain. Herein, samples of 25 µL of eight different concentrations of BKR1 peptide were distributed horizontally on 96-well plates, and each column had the same volume and the same concentration of the peptide. On the other hand, 25 mL of six different concentrations of a single antibiotic was added to different rows. Each row had the same concentration of the antibiotic. Meanwhile, two columns were reserved for testing the growth and negative controls. On each well, a fresh aliquot of 50 µL of bacterial suspension (106 CFU/mL) was added. The plates were then incubated overnight (18 h), at 37 °C, and the MIC values were then measured by reading the absorbance at λ = 600 nm throygh an ELISA plate reader (EpochTM; BioTek, Winooski, VT, USA).
The act of synergism was measured by calculating the Fractional Inhibitory Concentration Index (FICI) according to the following equation:
The results were then interpreted as synergistic if FIC value was £ 0.5, additive if FIC was in the range between 0.5–1, and indifferent if FIC was higher than 1 and lower than 4 (
Hemolytic assay
2 mL of human blood (Sigma Aldrich, St. Louis, MO, USA) was added to 48 mL of sterile phosphate buffer saline (PBS) at pH 7.4. The suspension was centrifuged three times at 2000 rpm for 5 min. Each time the supernatant was discarded and replaced by a fresh PBS buffer. The final concentration of human blood (RBCs) suspension was 4%. Eight tubes were then prepared, in which six of them contained 2 mL RBCs suspension and 2 mL of 6 different peptide concentration, one of them was for the positive control (2 mL of RBCs suspension mixed with 5µL of 0.1% Triton X-100), and the final tube represents the negative control (2 mL of RBCs suspension). Subsequently, all tubes were incubated at 37 °C for 60 min, then centrifuged at 2000 rpm for 5 min. After that, 1 mL was taken from each tube, and their absorbance was checked using (ELISA) microplate reader (EpochTM; BioTek, Winooski, VT, USA) at λ = 450 nm.
Cell culture
The cell lines used in the present study were Vero cells (ATCC CCL-81). The Vero lineage was isolated from kidney epithelial cells extracted from an African green monkey. The cells were grown in an RPMI media that consist of 10% fetal bovine serum and 1% v/v antibiotics (ampicillin, streptomycin), and antifungal agent (Amphotericin B), which were all obtained from Sigma-Aldrich (St. Louis, MO, USA). Cells were seeded in 75 cm² flasks with 24 mL of prepared media, incubated in 5% CO2 incubator at 37 °C. The media was changed every 24 h until the confluence reached 70%. The media was then discarded, leaving the adherent cells in the flask. A volume of 5–7 mL of trypsin 1X was added, and the flask was then returned to the CO2 incubator for 5–10 min. The trypsin was removed, and the detachment of the cells was confirmed through the use of an an inverted microscope. The trypsinization process was repeated until sufficient cell detachment. Finally, 5 mL media was added to neutralize the low pH of trypsin, followed by centrifugation at 2500 rpm for 5 min. The supernatant was discarded and replaced with 10 mL media and mixed by a vortex.
Cells were mixed with an equal amount of trypan blue (4%). The cells that were stained by trypan blue stain were considered dying cells, whereas live cells looked like stars. The cells were then counted using hemocytometer.
MTT cell proliferation assay
This assay was performed by seeding the cells at 5 × 105 cells/well in a 96-well plate flat bottom. The plates were incubated in a CO2 incubator at 37 °C for 18 h, to reattached cells to the bottom. The media was then discarded from the plates, and six different concentrations of peptide were dissolved in prepared RPMI media. The plates were incubated again in a CO2 incubator at 37 °C for 18 -24 hours. A volume of 25 µL of MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) suspension (4 mg/mL) was added, and the plates were further incubated for 4–6 hours. The MTT-Peptide solution was then removed, and 100 µL of DMSO was placed in each well followed by continuous pipetting to dissolve the formed Formazan crystals. The absorbance was then measured using ELISA microplate (EpochTM; BioTek, Winooski, VT, USA) at λ = 540 nm (
Results
Peptide design, synthesis, molecular modelling and In silico analysis
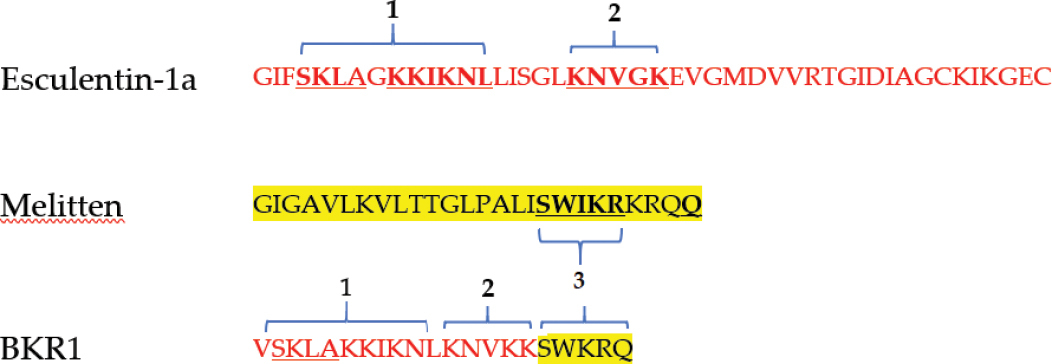
The sequence of the peptide was designed by fragmenting the helical parts of the parent peptides; Esculentin-1a and Melitten with some modifications as shown in Table
Prediction of secondary structure for the parent peptides and the hybrid peptide (BKR1) using Hierarchical Neural Network software (HNN).
| Name of Peptide | Sequence | # of a.a1 | α- helix % | b- sheet % | Random coils % |
|---|---|---|---|---|---|
|
46 | 45.00 | 19.00 | 34.78 | |
| 26 | 42.31 | 26.92 | 30.77 | ||
| 21 | 85.71 | 0.00 | 14.29 | ||

The physicochemical properties of BKR1 mentioned above were tested by ProtParam/ExPASy server and the Antimicrobial Peptide Database (APD3). The peptide is expected to display a stable form in the test tube as suggested by the instability index (>40), and the aliphatic index which provides an indication of the peptide’s thermostability. The peptide is considered hydrophilic (33% hydrophobic ratio) with high protein interaction ability (Boman index 2.81 kcal/mol). The charge of BKR1 peptide is +9 compared with +5 and +6 for Esculentin-1a and Melitten, respectively (Table
The physicochemical properties of the parent peptides and BKR1 peptide calculated by ProtParam from ExPASY and APD3.
| Peptide | MWT | pI | Instability index | Aliphatic index | Hydrophobic ratio | Total net charge | Boman index (kcal/mol) |
|---|---|---|---|---|---|---|---|
| Esculentin-1a | 4802.82 | 9.63 | -8.14 | 114.35 | 43% | 5 | 0.62 |
| Melitten | 2847.49 | 12.02 | 44.73 | 135.00 | 46% | 6 | 0.57 |
| BKR1 | 2525.31 | 11.51 | 16.68 | 88.10 | 33% | 9 | 2.81 |
BKR1 displays a net positive charge of +3 and a molecular weight of 1144.34 Da. BKR1 was purified to >98% purity using RP-HPLC (Suppl. material

Bacterial susceptibility assay
The minimal inhibitory concentrations (MIC) and the minimal bactericidal concentrations (MBC) for BKR1 displayed high antimicrobial activity against different Gram-negative bacteria. The average MIC value for all strains in this study was in the range of 25–30 mM, against the control strains and the resistant strains of the bacterial strains employed in the study (Table
Synergistic checkerboard assay
BKR1 peptide displayed significant antimicrobial activity against different strains of Gram-negative bacteria as shown in the previous section. However, we tested its potential of BKR1 to act as an antimicrobial adjuvant with the four suggested antibiotics (levofloxacin, LVX; chloramphenicol, CHL; ampicillin, AMP; and rifampicin, RIF) against different strains of P. aeruginosa (control strain ATCC 27853 and a resistant strain ATCC BAA-2114) in addition to E. coli (control strain ATCC 25922 and resistant strain ATCC BAA-2452). The act of synergism was checked using Checkerboard assay, and the results are summarized in Table
Summation of Fractional inhibitory concentration indices (FICI) for the synergistic Checkerboard Assay. LVX, levofloxacin; CHL, chloramphenicol; AMP, ampicillin; RIF, rifampicin; C.S., control strain; R.S. resistant strain; The percentage reduction in MIC was calculated using equation (1).
| Bacterial strain | Antibiotic | Peptide (BKR1) | FICI /Synergism | ||||||
|---|---|---|---|---|---|---|---|---|---|
| MIC alone (mM) | MIC in combination (mM) | Percentage reduction in MIC | MIC alone (mM) | MIC in combination (mM) | Percentage reduction in MIC | ||||
| P. aeruginosa C.S. (27853) | LVX | 0.25 | 0.083 | 67 | 25 | 6.25 | 75 | 0.58 | Additive |
| CHL | 25 | 5.25 | 79 | 25 | 3.75 | 85 | 0.36 | Synergistic | |
| AMP | 12.5 | 7.75 | 38 | 25 | 10 | 60 | 1.02 | Indifferent | |
| RIF | 15 | 2.5 | 83 | 25 | 2.5 | 90 | 0.27 | Synergistic | |
| P. aeruginosa R.S. (BAA-2114) | LVX | 12 | 3 | 75 | 25 | 1.25 | 95 | 0.30 | Synergistic |
| CHL | 200 | 25 | 88 | 25 | 1.25 | 95 | 0.18 | Synergistic | |
| AMP | >500 | 187.5 | >63 | 25 | 3.75 | 85 | >0.53 | Additive | |
| RIF | 50 | 5 | 90 | 25 | 2.5 | 90 | 0.20 | Synergistic | |
| E. coli C.S. (25922) | LVX | 0.25 | 0.188 | 25 | 25 | 2.5 | 90 | 0.85 | Additive |
| CHL | 10 | 5 | 50 | 25 | 1.25 | 95 | 0.55 | Additive | |
| AMP | 12.5 | 10 | 20 | 25 | 10 | 60 | 1.20 | Indifferent | |
| RIF | 8 | 2.5 | 69 | 25 | 1.25 | 95 | 0.36 | Synergistic | |
| E. coli R.S. (BAA-2452) | LVX | 25 | 3.75 | 85 | 25 | 2.5 | 90 | 0.25 | Synergistic |
| CHL | 25 | 5 | 80 | 25 | 2.5 | 90 | 0.30 | Synergistic | |
| AMP | >500 | 200 | >60 | 25 | 6.25 | 75 | >0.65 | – | |
| RIF | 5 | 1.25 | 75 | 25 | 1.25 | 95 | 0.30 | Synergistic | |
To calculate the percentage reduction in MIC, the following equation was used:
Interestingly, the combination of LVX/BKR1 displayed additive activity against control strains of both P. aeruginosa and E. coli but expressed synergistic activities against the resistant strains. The synergism of this combination against the tested strains caused a huge reduction in the MIC values of the combined agents (LVX: BKR1 = > 75%: > 90%).
CHL/BKR1 combination, on the other hand, displayed synergism with all tested strains with ≥ 80% reduction in the CHL MIC values and ≥ 85% in the MIC values of BKR1 (except against the control strain of E. coli, which showed additive effect). Whereas the combination of RIF/BKR1 always displayed strong synergism with more than 70% reduction in the MIC of both antibiotics, and more than 90% reduction in the MIC of BKR1 in all cases. AMP/BKR1 combination showed no synergism, but the outputs showed that their combinations decreased the MIC values for both agents in all cases as shown in Table
Hemolytic assay
The toxicity of antimicrobial peptides against human red blood cells has been a significant obstacle against their development into clinically useful drugs (

Mammalian cell cytotoxicity assay
The MTT assay evaluated the toxicity profile of BKR1 peptide against Vero cell line. The selectivity of the new antimicrobial agent towards bacteria is a major issue when designing novel antimicrobial agents. Therefore, different concentrations of the hybrid peptide were tested, and the outputs are represented in Fig.

Discussion
One of the major issues facing public health during last decade is the sustained emergence of bacterial resistance (
During the 21st century, several reports of the emergence of different multi-drug resistant bacteria (MDRB) were reported all around the world (
Antimicrobial peptides (AMPs) represents promising potential candidates for treating microbial infections (
In this work, we report the design and antimicrobial activity of a novel hybrid peptide (BKR1) based on two natural AMPs; Esculentin-1a and melittin. These peptides were chosen because they were cationic and displayed significant antimicrobial activity against Gram-negative bacteria (
The design strategy employed first melittin for the process of hybridization as this peptide is an attractive antimicrobial agent of 26 residues (
The novel hybrid peptide, BKR1 is a 21 amino acids peptide that was designed by joining truncated fragments from the N-terminal region of esculentin-1a and a modified part of the C-terminal of melittin. The MIC of BKR1 peptide was 25 mM against all tested strains with this study. The potency of the peptide against tested bacteria was the same regardless of the resistance degree. The MIC values of the parent peptides were reported in the literature; the MIC of melittin alone was recorded higher than 22.5 mM and 5.6 mM against P. aeruginosa (ATCC 27853) and E. coli (ATCC 25922), respectively (
Melittin causes 100% lysis of erythrocytes at only 10 mM (
Conclusion
In conclusion, we report the design of a novel hybrid peptide named BKR1 based on the amino acid sequence of Esculentin-1a and melittin. The hybrid peptide displayed strong antimicrobial activities against planktonic Gram-negative bacterial strains including resistant clinical isolates. BKR1 displayed bactericidal activity and showed similar potency against bacteria regardless of its resistance degree. The hemolytic activity of BKR1 at its antimicrobial concentrations were negligible however, it still risks the viability of the mammalian cells as indicated in the cell proliferation assays. The synergism studies were performed to decrease the MIC values of the peptide to make it safer to use in mammalian cells, and to decrease any possibilities for future resistance against this peptide. The synergism was the strongest with chloramphenicol and rifampicin against P. aeruginosa resistant strain, even though these antibiotics are not effective when used alone. On the other hand, the synergism with levofloxacin was the strongest against the E. coli resistant strain, regardless of its resistance against this antibiotic when used alone.
Acknowledgements
The authors express their gratitude to Dr. Rawan Alsharideh and Dr. Mohammad Alsaggar for their valuable comments. This project was funded by the Deanship of Research at the Jordan University of Science and Technology, grant number 592-2018.
References
- Alekshun MN, Levy SB (2007) Molecular mechanisms of antibacterial multidrug resistance. Cell 128(6): 1037–1050. https://doi.org/10.1016/j.cell.2007.03.004
- Ahmed TAE, Hammami R (2019) Recent insights into structure-function relationships of antimicrobial peptides. Journal of Food Biochemistry 43(1): e12546. https://doi.org/10.1111/jfbc.12546
- Almaaytah A, Tarazi S, Abu-Alhaijaa A, Altall Y, Alshar’i N, Bodoor K, Al-Balas Q (2014) Enhanced antimicrobial activity of AamAP1-Lysine, a novel synthetic peptide analog derived from the scorpion venom peptide AamAP1. Pharmaceuticals (Basel, Switzerland) 7(5): 502–516. https://doi.org/10.3390/ph7050502
- Almaaytah A, Qaoud MT, Abualhaijaa A, Al-Balas Q, Alzoubi KH (2018) Hybridization and antibiotic synergism as a tool for reducing the cytotoxicity of antimicrobial peptides. Infection and Drug Resistance 11: 835–847. https://doi.org/10.2147/IDR.S166236
- Almaaytah A, Abualhaijaa A, Alqudah O (2019) The evaluation of the synergistic antimicrobial and antibiofilm activity of AamAP1-Lysine with conventional antibiotics against representative resistant strains of both Gram-positive and Gram-negative bacteria. Infection and Drug Resistance 12: 1371–1380. https://doi.org/10.2147/IDR.S204626
- Al Tall Y, Abualhaijaa A, Alsaggar M, Almaaytah A, Masadeh M, Alzoubi KH (2019) Design and characterization of a new hybrid peptide from LL-37 and BMAP-27. Infection and Drug Resistance 12: 1035–1045. https://doi.org/10.2147/IDR.S199473
- Arias CA, Murray BE (2009) Antibiotic-resistant bugs in the 21st century--a clinical super-challenge. The New England Journal of Medicine 360(5): 439–443. https://doi.org/10.1056/NEJMp0804651
- Asthana N, Yadav SP, Ghosh JK (2004) Dissection of antibacterial and toxic activity of melittin: a leucine zipper motif plays a crucial role in determining its hemolytic activity but not antibacterial activity. The Journal of Biological Chemistry 279(53): 55042–55050. https://doi.org/10.1074/jbc.M408881200
- Bassetti M, Ginocchio F, Mikulska M (2011) New treatment options against gram-negative organisms. Critical Care (London, England) 15(2): 215. https://doi.org/10.1186/cc9997
- Condie CC, Quay SC (1983) Conformational studies of aqueous melittin. Characteristics of a fluorescent probe binding site. The Journal of Biological Chemistry 258(13): 8231–8234. https://doi.org/10.1016/S0021-9258(20)82053-5
- Conlon JM, Al-Kharrge R, Ahmed E, Raza H, Galadari S, Condamine E (2007) Effect of aminoisobutyric acid (Aib) substitutions on the antimicrobial and cytolytic activities of the frog skin peptide, temporin-1DRa. Peptides 28(10): 2075–2080. https://doi.org/10.1016/j.peptides.2007.07.023
- Daikos GL, Petrikkos P, Psichogiou M, Kosmidis C, Vryonis E, Skoutelis A, Georgousi K, Tzouvelekis LS, Tassios PT, Bamia C, Petrikkos G (2009) Prospective observational study of the impact of VIM-1 metallo-beta-lactamase on the outcome of patients with Klebsiella pneumoniae bloodstream infections. Antimicrobial Agents and Chemotherapy 53(5): 1868–1873. https://doi.org/10.1128/AAC.00782-08
- Desbois AP, Gemmell CG, Coote PJ (2010) In vivo efficacy of the antimicrobial peptide ranalexin in combination with the endopeptidase lysostaphin against wound and systemic meticillin-resistant Staphylococcus aureus (MRSA) infections. International Journal of Antimicrobial Agents 35(6): 559–565. https://doi.org/10.1016/j.ijantimicag.2010.01.016
- Di Grazia A, Cappiello F, Imanishi A, Mastrofrancesco A, Picardo M, Paus R, Mangoni ML (2015) The frog skin-derived antimicrobial peptide Esculentin-1a(1-21)NH2 promotes the migration of human HaCaT keratinocytes in an EGF receptor-dependent manner: A novel promoter of human skin wound healing?. PloS ONE 10(6): e0128663. https://doi.org/10.1371/journal.pone.0128663
- Di Grazia A, Cappiello F, Cohen H, Casciaro B, Luca V, Pini A, Di YP, Shai Y, Mangoni ML (2015) D-Amino acids incorporation in the frog skin-derived peptide esculentin-1a(1-21)NH2 is beneficial for its multiple functions. Amino Acids 47(12): 2505–2519. https://doi.org/10.1007/s00726-015-2041-y
- Dosler S, Karaaslan E, Alev Gerceker A (2016) Antibacterial and anti-biofilm activities of melittin and colistin, alone and in combination with antibiotics against Gram-negative bacteria. Journal of Chemotherapy (Florence, Italy) 28(2): 95–103. https://doi.org/10.1179/1973947815Y.0000000004
- Erdem Büyükkiraz M, Kesmen Z (2022) Antimicrobial peptides (AMPs): A promising class of antimicrobial compounds. Journal of Applied Microbiology 132(3): 1573–1596. https://doi.org/10.1111/jam.15314
- Fetar H, Gilmour C, Klinoski R, Daigle DM, Dean CR, Poole K (2011) mexEF-oprN multidrug efflux operon of Pseudomonas aeruginosa: regulation by the MexT activator in response to nitrosative stress and chloramphenicol. Antimicrobial Agents and Chemotherapy 55(2): 508–514. https://doi.org/10.1128/AAC.00830-10
- Gabere MN, Noble WS (2017) Empirical comparison of web-based antimicrobial peptide prediction tools. Bioinformatics (Oxford, England) 33(13): 1921–1929. https://doi.org/10.1093/bioinformatics/btx081
- Garnacho-Montero J, Sa-Borges M, Sole-Violan J, Barcenilla F, Escoresca-Ortega A, Ochoa M, Cayuela A, Rello J (2007) Optimal management therapy for Pseudomonas aeruginosa ventilator-associated pneumonia: an observational, multicenter study comparing monotherapy with combination antibiotic therapy. Critical Care Medicine 35(8): 1888–1895. https://doi.org/10.1097/01.CCM.0000275389.31974.22
- Geitani R, Ayoub Moubareck C, Touqui L, Karam Sarkis D (2019) Cationic antimicrobial peptides: alternatives and/or adjuvants to antibiotics active against methicillin-resistant Staphylococcus aureus and multidrug-resistant Pseudomonas aeruginosa. BMC microbiology 19(1): 54. https://doi.org/10.1186/s12866-019-1416-8
- Ghisalberti D, Masi M, Pagès JM, Chevalier J (2005) Chloramphenicol and expression of multidrug efflux pump in Enterobacter aerogenes. Biochemical and Biophysical Research Communications 328(4): 1113–1118. https://doi.org/10.1016/j.bbrc.2005.01.069
- Giacometti A, Cirioni O, Barchiesi F, Scalise G (2000) In-vitro activity and killing effect of polycationic peptides on methicillin-resistant Staphylococcus aureus and interactions with clinically used antibiotics. Diagnostic Microbiology and Infectious Disease 38(2): 115–118. https://doi.org/10.1016/s0732-8893(00)00175-9
- Jasovský D, Littmann J, Zorzet A, Cars O (2016) Antimicrobial resistance-a threat to the world’s sustainable development. Upsala Journal of Medical Sciences 121(3): 159–164. https://doi.org/10.1080/03009734.2016.1195900
- Jiang Z, Vasil AI, Hale JD, Hancock RE, Vasil ML, Hodges RS (2008) Effects of net charge and the number of positively charged residues on the biological activity of amphipathic alpha-helical cationic antimicrobial peptides. Biopolymers 90(3): 369–383. https://doi.org/10.1002/bip.20911
- Khabbaz RF, Moseley RR, Steiner RJ, Levitt AM, Bell BP (2014) Challenges of infectious diseases in the USA. Lancet (London, England) 384(9937): 53–63. https://doi.org/10.1016/S0140-6736(14)60890-4
- Labby KJ, Garneau-Tsodikova S (2013) Strategies to overcome the action of aminoglycoside-modifying enzymes for treating resistant bacterial infections. Future Medicinal Chemistry 5(11): 1285–1309. https://doi.org/10.4155/fmc.13.80
- Lerminiaux NA, Cameron ADS (2019) Horizontal transfer of antibiotic resistance genes in clinical environments. Canadian Journal of Microbiology 65(1): 34–44. https://doi.org/10.1139/cjm-2018-0275
- Lata S, Sharma BK, Raghava GP (2007) Analysis and prediction of antibacterial peptides. BMC bioinformatics 8: 263. https://doi.org/10.1186/1471-2105-8-263
- Leveritt III JM, Pino-Angeles A, Lazaridis T (2015) The structure of a melittin-stabilized pore. Biophysical Journal 108(10): 2424–2426. https://doi.org/10.1016/j.bpj.2015.04.006
- Levy SB (2005) Antibiotic resistance-the problem intensifies. Advanced Drug Delivery Reviews 57(10): 1446–1450. https://doi.org/10.1016/j.addr.2005.04.001
- Lovell SC, Davis IW, Arendall III WB, de Bakker PI, Word JM, Prisant MG, Richardson JS, Richardson DC (2003) Structure validation by Calpha geometry: phi, psi and Cbeta deviation. Proteins 50(3): 437–450. https://doi.org/10.1002/prot.10286
- Luna-Ramirez K, Tonk M, Rahnamaeian M, Vilcinskas A (2017) Bioactivity of natural and engineered antimicrobial peptides from venom of the scorpions Urodacus yaschenkoi and U. manicatus. Toxins 9(1): 22. https://doi.org/10.3390/toxins9010022
- Mangoni ML, Maisetta G, Di Luca M, Gaddi LM, Esin S, Florio W, Brancatisano FL, Barra D, Campa M, Batoni G (2008) Comparative analysis of the bactericidal activities of amphibian peptide analogues against multidrug-resistant nosocomial bacterial strains. Antimicrobial Agents and Chemotherapy 52(1): 85–91. https://doi.org/10.1128/AAC.00796-07
- Melander RJ, Melander C (2017) The challenge of overcoming antibiotic resistance: An adjuvant approach?. ACS Infectious Diseases 3(8): 559–563. https://doi.org/10.1021/acsinfecdis.7b00071
- Mohseni S, Aghayan M, Ghorani-Azam A, Behdani M, Asoodeh A (2015) Evaluation of antibacterial properties of Barium Zirconate Titanate (BZT) nanoparticle. Brazilian Journal of Microbiology: [publication of the Brazilian Society for Microbiology] 45(4): 1393–1399. https://doi.org/10.1590/s1517-83822014000400033
- Morens DM, Fauci AS (2012) Emerging infectious diseases in 2012: 20 years after the institute of medicine report. mBio 3(6): e00494-12. https://doi.org/10.1128/mBio.00494-12
- Morrison L, Zembower TR (2020) Antimicrobial resistance. Gastrointestinal Endoscopy Clinics of North America 30(4): 619–635. https://doi.org/10.1016/j.giec.2020.06.004
- Omran BA, Nassar HN, Fatthallah NA, Hamdy A, El-Shatoury EH, El-Gendy NS (2018) Characterization and antimicrobial activity of silver nanoparticles mycosynthesized by Aspergillus brasiliensis. Journal of Applied Microbiology 125(2): 370–382. https://doi.org/10.1111/jam.13776
- Oren Z, Shai Y (1997) Selective lysis of bacteria but not mammalian cells by diastereomers of melittin: structure-function study. Biochemistry 36(7): 1826–1835. https://doi.org/10.1021/bi962507l
- Ostroumova OS, Efimova SS, Malev VV (2015) Modifiers of membrane dipole potentials as tools for investigating ion channel formation and functioning. International Review of Cell and Molecular Biology 315: 245–297. https://doi.org/10.1016/bs.ircmb.2014.12.001
- Pasupuleti M, Schmidtchen A, Malmsten M (2012) Antimicrobial peptides: key components of the innate immune system. Critical Reviews in Biotechnology 32(2): 143–171. https://doi.org/10.3109/07388551.2011.594423
- Petrosillo N, Ioannidou E, Falagas ME (2008) Colistin monotherapy vs. combination therapy: evidence from microbiological, animal and clinical studies. Clinical Microbiology and Infection: the official publication of the European Society of Clinical Microbiology and Infectious Diseases 14(9): 816–827. https://doi.org/10.1111/j.1469-0691.2008.02061.x
- Pouny Y, Rapaport D, Mor A, Nicolas P, Shai Y (1992) Interaction of antimicrobial dermaseptin and its fluorescently labeled analogues with phospholipid membranes. Biochemistry 31(49): 12416–12423. https://doi.org/10.1021/bi00164a017
- Pfalzgraff A, Brandenburg K, Weindl G (2018) Antimicrobial peptides and their therapeutic potential for bacterial skin infections and wounds. Frontiers in Pharmacology 9: 281. https://doi.org/10.3389/fphar.2018.00281
- Samonis G, Maraki S, Karageorgopoulos DE, Vouloumanou EK, Falagas ME (2012) Synergy of fosfomycin with carbapenems, colistin, netilmicin, and tigecycline against multidrug-resistant Klebsiella pneumoniae, Escherichia coli, and Pseudomonas aeruginosa clinical isolates. European Journal of Clinical Microbiology & Infectious Diseases: official publication of the European Society of Clinical Microbiology 31(5): 695–701. https://doi.org/10.1007/s10096-011-1360-5
- Schmidt NW, Wong GC (2013) Antimicrobial peptides and induced membrane curvature: geometry, coordination chemistry, and molecular engineering. Current opinion in solid state & materials science 17(4): 151–163. https://doi.org/10.1016/j.cossms.2013.09.004
- Seo MD, Won HS, Kim JH, Mishig-Ochir T, Lee BJ (2012) Antimicrobial peptides for therapeutic applications: a review. Molecules (Basel, Switzerland) 17(10): 12276–12286. https://doi.org/10.3390/molecules171012276
- Sheikholeslami S, Mousavi SE, Ahmadi Ashtiani HR, Hosseini Doust SR, Mahdi Rezayat S (2016) Antibacterial activity of silver nanoparticles and their combination with Zataria multiflora essential oil and methanol extract. Jundishapur Journal of Microbiology 9(10): e36070. https://doi.org/10.5812/jjm.36070
- Simmaco M, Mignogna G, Barra D, Bossa F (1993) Novel antimicrobial peptides from skin secretion of the European frog Rana esculenta. FEBS Letters 324(2): 159–161. https://doi.org/10.1016/0014-5793(93)81384-c
- Simmaco M, Mignogna G, Barra D (1998) Antimicrobial peptides from amphibian skin: what do they tell us? Biopolymers 47(6): 435–450. https://doi.org/10.1002/(SICI)1097-0282(1998)47:6<435::AID-BIP3>3.0.CO;2-8
- Sun J, Deng Z, Yan A (2014) Bacterial multidrug efflux pumps: mechanisms, physiology and pharmacological exploitations. Biochemical and Biophysical Research Communications 453(2): 254–267. https://doi.org/10.1016/j.bbrc.2014.05.090
- Tender T, Rahangdale RR, Balireddy S, Nampoothiri M, Sharma KK, Raghu Chandrashekar H (2021) Melittin, a honeybee venom derived peptide for the treatment of chemotherapy-induced peripheral neuropathy. Medical Oncology (Northwood, London, England) 38(5): 52. https://doi.org/10.1007/s12032-021-01496-9
- Waldman EA, Sato AP (2016) Path of infectious diseases in Brazil in the last 50 years: an ongoing challenge. Revista de Saude Publica 50: 68. https://doi.org/10.1590/S1518-8787.2016050000232
- Wang G, Li X, Wang Z (2016) APD3: the antimicrobial peptide database as a tool for research and education. Nucleic Acids Research 44(D1): D1087–D1093. https://doi.org/10.1093/nar/gkv1278
- Webb B, Sali A (2016) Comparative protein structure modeling using MODELLER. Current Protocols in Bioinformatics 54: 5.6.1–5.6.37. https://doi.org/10.1002/cpbi.3
- Wilkins MR, Gasteiger E, Bairoch A, Sanchez JC, Williams KL, Appel RD, Hochstrasser DF (1999) Protein identification and analysis tools in the ExPASy server. Methods in Molecular Biology (Clifton, N.J. ) 112: 531–552. https://doi.org/10.1385/1-59259-584-7:531
- Wimley WC (2010) Describing the mechanism of antimicrobial peptide action with the interfacial activity model. ACS Chemical Biology 5(10): 905–917. https://doi.org/10.1021/cb1001558
- Worthington RJ, Melander C (2013) Combination approaches to combat multidrug-resistant bacteria. Trends in biotechnology 31(3): 177–184. https://doi.org/10.1016/j.tibtech.2012.12.006
- Wright GD (2016) Antibiotic adjuvants: Rescuing antibiotics from resistance. Trends in Microbiology 24(11): 862–871. https://doi.org/10.1016/j.tim.2016.06.009
- Yeung AT, Gellatly SL, Hancock RE (2011) Multifunctional cationic host defence peptides and their clinical applications. Cellular and Molecular Life Sciences: CMLS 68(13): 2161–2176. https://doi.org/10.1007/s00018-011-0710-x
- Zasloff M (2002) Antimicrobial peptides of multicellular organisms. Nature 415(6870): 389–395. https://doi.org/10.1038/415389a
- Zhang Y (2008) I-TASSER server for protein 3D structure prediction. BMC bioinformatics 9: 40. https://doi.org/10.1186/1471-2105-9-40
- Zhang Y, Liu Y, Sun Y, Liu Q, Wang X, Li Z, Hao J (2014) In vitro synergistic activities of antimicrobial peptide brevinin-2CE with five kinds of antibiotics against multidrug-resistant clinical isolates. Current Microbiology 68(6): 685–692. https://doi.org/10.1007/s00284-014-0529-4
- Zimmermann L, Stephens A, Nam SZ, Rau D, Kübler J, Lozajic M, Gabler F, Söding J, Lupas AN, Alva V (2018) A completely reimplemented MPI bioinformatics toolkit with a new HHpred server at its core. Journal of Molecular Biology 430(15): 2237–2243. https://doi.org/10.1016/j.jmb.2017.12.007
Supplementary material
RP-HPLC analysis Chromatogram of BKR1 peptide
Data type: figure (word document).