|
Corresponding author: Pavlina Andreeva-Gateva ( pandreeva_gateva@outlook.com ) Academic editor: Georgi Momekov
© 2020 Pavlina Andreeva-Gateva, Dimitar Bakalov, Zafer Sabit, Radka Tafradjiiska-Hadjiolova.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Andreeva-Gateva P, Bakalov D, Sabit Z, Tafradjiiska-Hadjiolova R (2020) Aryl hydrocarbon receptors as potential therapeutic targets. Pharmacia 67(4): 311-315. https://doi.org/10.3897/pharmacia.67.e47298
|
Abstract
Aryl hydrocarbon receptors (AhR) are regulators of the expression of cytochrome P-450 isoforms, mediating a wide variety of the effects of substances from the endogenous or exogenous origin, including those produced from the microbiome. An exciting new aspect of their activity is their localization in the brain and their potential to modulate the action of the immune system. AhR is emerging as an essential toxicological and therapeutic target for neuromodulation. Further studies are needed for elucidating their utility as drug-targets.
Keywords
dioxin, indoles, toxicology, neuroprotection
Introduction
Aryl hydrocarbon receptors (AhR) are ligand-activated receptors. They form nuclear heterodimer complexes with AhR-dependent nuclear translocator protein, and this complex binds to cis-xenobiotic responsive elements in the promoter region of AhR-responsive genes (
Many of those ligands have a much lower binding affinity for AhR than TCDD and structure-like toxic halogenated aromatic substances (
After being identified as mediators of the cellular response to xenobiotics, epidemiological studies in humans were conducted. The attention was focused on the pathological conditions of the immune system, lipid metabolism, epithelial integrity, porphyria manifestations, thymus involution, and neoplasms. The involvement of AhR inspired scientific researches (
| Chemical structure and name | Source | References | Chemical structure and name | Source | References |
|---|---|---|---|---|---|
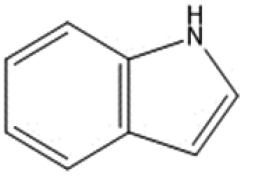
|
microbial | Hubbard (2015) |
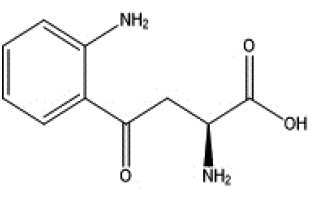
|
endogenous |
|
| Indole | Kynurenine | ||||
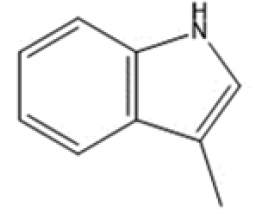
|
microbial | Hubbard (2015) |
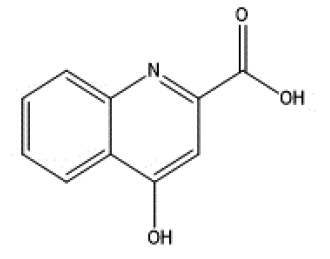
|
endogenous |
|
| Skatole | Kynurenic acid | ||||
|
microbial | Hubbard (2015) |
|
endogenous |
|
| 3-hydroxy indole | Xanthurenic acid | ||||
|
microbial endogenous |
|
|
endogenous |
|
| Indole-3-acetaldehyde | Cinnabarinic acid | ||||
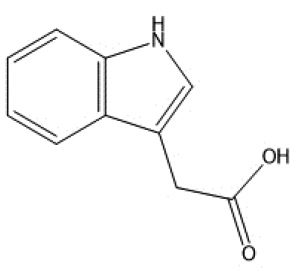
|
microbial endogenous |
|
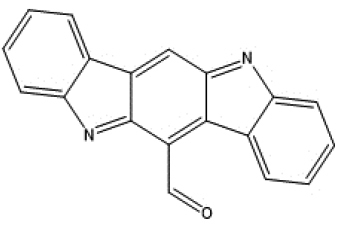
|
endogenous |
|
| Indole-3-acetic acid | FICZ | ||||
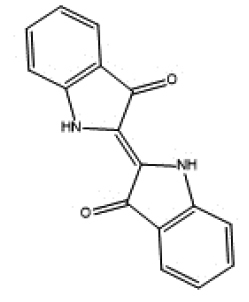
|
microbial |
|
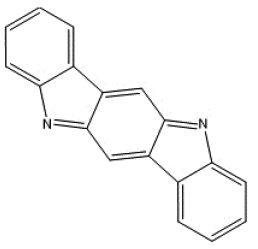
|
endogenous |
|
| Indigo | Indolo(3,2-b) carbazole | ||||
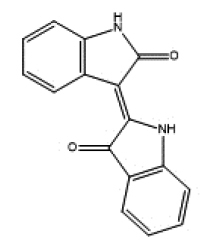
|
microbial | Flaveny et al. (2009) |
|
dietary |
|
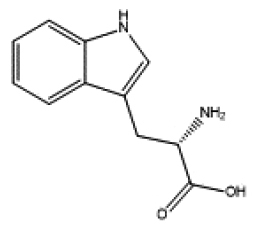
| indirubin | Tryptophan | ||||
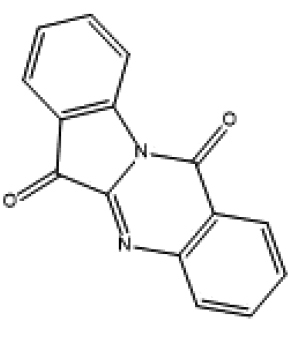
|
microbial |
|
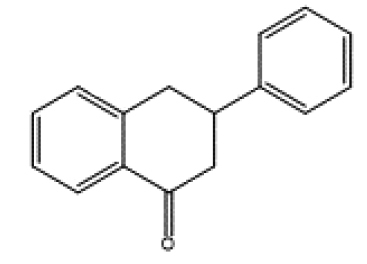
|
dietary |
|
| Truptanthrin | Flavonoids |
AhR as a therapeutic target
The identification of AhR ligands and their well-described positive health effect and beneficial pharmaceutical properties has stimulated studies aimed at developing drugs for various tumors, immune and inflammatory diseases, and enhancers of hematopoietic stem cell production. In the development of drugs that target AhR, the aim is mainly directed to selective AhR modulators, in which the ligand exhibits tissue-specific agonist or antagonist activity (
Since AhRs are widely expressed in a number of tumors, molecules with antagonistic activity against AhRs could be considered as potential candidates for the treatment of such diseases. The most well-known AhR antagonist is alpha-naphtoflavone (Gasievicz and Rucci 1991). The potent AhR antagonist StemRegenin 1 has recently been developed as an inducer of human hematopoietic stem cell proliferation in vitro (
Throughout the many plant nutrients and chemicals of plant origin in the human diet, flavonoids are the most abundant and ubiquitous in fruits, vegetables, and wine. Quercetin, apigenin, and campherol, which are included in some foods, such as rose hips, linden flowers, honey, grapes, have been shown to exert agonist / antagonistic effects on AhR in various tissues (
Neuroprotective properties of AhR modulators
AhR expression in vertebrate brain has recently been demonstrated by immunohistochemistry, with the brain stem, pineal gland, and some hypothalamic nuclei (including the suprachiasmatic nucleus controlling the circadian rhythm) having significantly elevated AhR levels compared to other areas of the brain (
Conclusion
All these data put the importance of AhR as a toxicological and pharmacological target. Further evaluation of the neuropharmacological potential of substances that bind and modulate AhR is needed.
Acknowledgements
With the support of the Council of Medical Sciences of the Medical University of Sofia, grant D-62/2019.
References
- Abdelrahim M, Ariazi E, Kim K, Khan S, Barhoumi R, Burghardt R, Liu S, Hill D, Finnell R, Wlodarczyk B, Jordan VC, Safe S (2006) 3-Methylcholanthrene and Other Aryl Hydrocarbon Receptor Agonists Directly Activate Estrogen Receptor α. Cancer Research 66(4): 2459–2467. https://doi.org/10.1158/0008-5472.CAN-05-3132
- Bock KW, Köhle C (2010) Contributions of the Ah receptor to bilirubin homeostasis and its antioxidative and atheroprotective functions. Biological Chemistry 391(6): 645–653. https://doi.org/10.1515/bc.2010.065
- Boitano AE, Wang J, Romeo R, Bouchez LC, Parker AE, Sutton SE, Walker JR, Flaveny CA, Perdew GH, Denison MS, Schultz PG, Cooke MP (2010) Aryl Hydrocarbon Receptor Antagonists Promote the Expansion of Human Hematopoietic Stem Cells. Science 329(5997): 1345–1348. https://doi.org/10.1126/science.1191536.
- Céspedes MA, Galindo MI, Couso JP (2010) Dioxin Toxicity In Vivo Results from an Increase in the Dioxin-Independent Transcriptional Activity of the Aryl Hydrocarbon Receptor. PLoS ONE 5(11): e15382. https://doi.org/10.1371/journal.pone.0015382
- Cuartero MI, Ballesteros I, de la Parra J, Harkin AL, Abautret-Daly A, Sherwin E, Fernández-Salguero P, Corbí ÁL, Lizasoain I, Moro MA (2014) L-Kynurenine/Aryl Hydrocarbon Receptor Pathway Mediates Brain Damage After Experimental Stroke. Circulation 130(23): 2040–2051. https://doi.org/10.1161/CIRCULATIONAHA.114.011394
- Denison MS, Heath-Pagliuso S (1998) The Ah Receptor: A Regulator of the Biochemical and Toxicological Actions of Structurally Diverse Chemicals. Bulletin of Environmental Contamination and Toxicology 61(5): 557–568. https://doi.org/10.1007/PL00002973
- Denison MS, Soshilov AA, He G, DeGroot DE, Zhao B (2011) Exactly the Same but Different: Promiscuity and Diversity in the Molecular Mechanisms of Action of the Aryl Hydrocarbon (Dioxin) Receptor. Toxicological Sciences 124(1): 1–22. https://doi.org/10.1093/toxsci/kfr218
- Flesch-Janys D, Berger J, Gum P, Manz A, Nagel S, Waltsgott H, Dwyer JH (1995) Exposure to Polychlorinated Dioxins and Furans (PCDD/F) and Mortality in a Cohort of Workers from a Herbicide-producing Plant in Hamburg, Federal Republic of Germany. American Journal of Epidemiology 142(11): 1165–1175. https://doi.org/10.1093/oxfordjournals.aje.a117575
- Gasiewicz TA, Rucci G (1991) Alpha-naphthoflavone acts as an antagonist of 2,3,7,8-tetrachlorodibenzo-p-dioxin by forming an inactive complex with the Ah receptor. Molecular Pharmacology 40(5): 607–612.
- Hooper LV (2011) You AhR What You Eat: Linking Diet and Immunity. Cell 147(3): 489–491. https://doi.org/10.1016/j.cell.2011.10.004
- Hubbard TD, Murray IA, Perdew GH (2015) Indole and Tryptophan Metabolism: Endogenous and Dietary Routes to Ah Receptor Activation. Drug Metabolism and Disposition 43(10): 1522–1535. https://doi.org/10.1124/dmd.115.064246
- Jin U-H, Kim S-B, Safe S (2015) Omeprazole Inhibits Pancreatic Cancer Cell Invasion through a Nongenomic Aryl Hydrocarbon Receptor Pathway. Chemical Research in Toxicology 28(5): 907–918. https://doi.org/10.1021/tx5005198
- Jin U-H, Lee S, Safe S (2012) Aryl Hydrocarbon Receptor (AHR)-Active Pharmaceuticals Are Selective AHR Modulators in MDA-MB-468 and BT474 Breast Cancer Cells. Journal of Pharmacology and Experimental Therapeutics 343(2): 333–341. https://doi.org/10.1124/jpet.112.195339
- Jin U-H, Lee S-O, Sridharan G, Lee K, Davidson LA, Jayaraman A, Chapkin RS, Alaniz R, Safe S (2014) Microbiome-Derived Tryptophan Metabolites and Their Aryl Hydrocarbon Receptor-Dependent Agonist and Antagonist Activities. Molecular Pharmacology 85(5): 777–788. https://doi.org/10.1124/mol.113.091165
- Juricek L, Coumoul X (2018) The Aryl Hydrocarbon Receptor and the Nervous System. International Journal of Molecular Sciences 19(9): e2504. https://doi.org/10.3390/ijms19092504
- Korecka A, Dona A, Lahiri S, Tett AJ, Al-Asmakh M, Braniste V, D’Arienzo R, Abbaspour A, Reichardt N, Fujii-Kuriyama Y, Rafter J, Narbad A, Holmes E, Nicholson J, Arulampalam V, Pettersson S (2016) Bidirectional communication between the Aryl hydrocarbon Receptor (AhR) and the microbiome tunes host metabolism. npj Biofilms and Microbiomes 2: e16014. https://doi.org/10.1038/npjbiofilms.2016.14
- Lahvis GP, Lindell SL, Thomas RS, McCuskey RS, Murphy C, Glover E, Bentz M, Southard J, Bradfield CA (2000) Portosystemic shunting and persistent fetal vascular structures in aryl hydrocarbon receptor-deficient mice. Proceedings of the National Academy of Sciences of the United States of America 97(19): 10442–10447. https://doi.org/10.1073/pnas.190256997
- Lin C-H, Juan S-H, Wang CY, Sun Y-Y, Chou C-M, Chang S-F, Hu S-Y, Lee W-S, Lee Y-H (2008) Neuronal activity enhances aryl hydrocarbon receptor-mediated gene expression and dioxin neurotoxicity in cortical neurons. Journal of Neurochemistry 104(5): 1415–1429. https://doi.org/10.1111/j.1471-4159.2007.05098.x
- Liu Y, Li P, Lu J, Xiong W, Oger J, Tetzlaff W, Cynader M (2008) Bilirubin Possesses Powerful Immunomodulatory Activity and Suppresses Experimental Autoimmune Encephalomyelitis. The Journal of Immunology 181(3): 1887–1897. https://doi.org/10.4049/jimmunol.181.3.1887
- Lowe MM, Mold JE, Kanwar B, Huang Y, Louie A, Pollastri MP, Wang C, Patel G, Franks DG, Schlezinger J, Sherr DH, Silverstone AE, Hahn ME, McCune JM (2014) Identification of Cinnabarinic Acid as a Novel Endogenous Aryl Hydrocarbon Receptor Ligand That Drives IL-22 Production. PLoS ONE 9: e87877. https://doi.org/10.1371/journal.pone.0087877
- Mulero-Navarro S, Fernandez-Salguero PM (2016) New Trends in Aryl Hydrocarbon Receptor Biology. Frontiers in Cell and Developmental Biology 4: e45. https://doi.org/10.3389/fcell.2016.00045
- Murray IA, Perdew GH (2017) Ligand activation of the Ah receptor contributes to gastrointestinal homeostasis. Current Opinion in Toxicology 2: 15–23. https://doi.org/10.1016/j.cotox.2017.01.003
- Novikov O, Wang Z, Stanford EA, Parks AJ, Ramirez-Cardenas A, Landesman E, Laklouk I, Sarita-Reyes C, Gusenleitner D, Li A, Monti S, Manteiga S, Lee K, Sherr DH (2016) An Aryl Hydrocarbon Receptor-Mediated Amplification Loop That Enforces Cell Migration in ER − /PR − /Her2 − Human Breast Cancer Cells. Molecular Pharmacology 90(5): 674–688. https://doi.org/10.1124/mol.116.105361
- Papoutsis AJ, Lamore SD, Wondrak GT, Selmin OI, Romagnolo DF (2010) Resveratrol Prevents Epigenetic Silencing of BRCA-1 by the Aromatic Hydrocarbon Receptor in Human Breast Cancer Cells. The Journal of Nutrition 140(9): 1607–1614. https://doi.org/10.3945/jn.110.123422
- Poland A, Glover E, Kende AS (1976) Stereospecific, high affinity binding of 2,3,7,8-tetrachlorodibenzo-p-dioxin by hepatic cytosol. Evidence that the binding species is receptor for induction of aryl hydrocarbon hydroxylase. The Journal of Biological Chemistry 251(16): 4936–4946.
- Qin H, Powell-Coffman JA (2004) The Caenorhabditis elegans aryl hydrocarbon receptor, AHR-1, regulates neuronal development. Developmental Biology 270(1): 64–75. https://doi.org/10.1016/j.ydbio.2004.02.004
- Reed GA (2005) A Phase I Study of Indole-3-Carbinol in Women: Tolerability and Effects. Cancer Epidemiology Biomarkers & Prevention 14(8): 1953–1960. https://doi.org/10.1158/1055-9965.EPI-05-0121
- Romagnolo DF, Daniels KD, Grunwald JT, Ramos SA, Propper CR, Selmin OI (2016) Epigenetics of breast cancer: Modifying role of environmental and bioactive food compounds. Molecular Nutrition & Food Research 60(6): 1310–1329. https://doi.org/10.1002/mnfr.201501063
- Safe S (2017) Carbidopa: a selective Ah receptor modulator (SAhRM). Biochemical Journal 474(22): 3763–3765. https://doi.org/10.1042/BCJ20170728
- Shertzer HG, Senft AP (2000) The Micronutrient Indole-3-Carbinol: Implications for Disease and Chemoprevention. Drug Metabolism and Personalized Therapy 17(1–4): 159–188. https://doi.org/10.1515/DMDI.2000.17.1-4.159
- Sugihara K, Kitamura S, Yamada T, Okayama T, Ohta S, Yamashita K, Yasuda M, Fujii-Kuriyama Y, Saeki K, Matsui S, Matsuda T (2004) Aryl hydrocarbon receptor-mediated induction of microsomal drug-metabolizing enzyme activity by indirubin and indigo. Biochemical and Biophysical Research Communications 318(2): 571–578. https://doi.org/10.1016/j.bbrc.2004.04.066
- Wan C, Zhang Y, Jiang J, Jiang S, Nie X, Li A, Guo A, Wu Q (2015) Critical Role of TAK1-Dependent Nuclear Factor-κB Signaling in 2,3,7,8-Tetrachlorodibenzo-p-dioxin-induced Astrocyte Activation and Subsequent Neuronal Death. Neurochemical Research 40: 1220–1231. https://doi.org/10.1007/s11064-015-1585-2
- Wincent E, Amini N, Luecke S, Glatt H, Bergman J, Crescenzi C, Rannug A, Rannug U (2009) The Suggested Physiologic Aryl Hydrocarbon Receptor Activator and Cytochrome P4501 Substrate 6-Formylindolo[3,2- b] carbazole Is Present in Humans. Journal of Biological Chemistry 284: 2690–2696. https://doi.org/10.1074/jbc.M808321200
- Zelante T, Iannitti RG, Cunha C, De Luca A, Giovannini G, Pieraccini G, Zecchi R, D’Angelo C, Massi-Benedetti C, Fallarino F, Carvalho A, Puccetti P, Romani L (2013) Tryptophan Catabolites from Microbiota Engage Aryl Hydrocarbon Receptor and Balance Mucosal Reactivity via Interleukin-22. Immunity 39(2): 372–385. https://doi.org/10.1016/j.immuni.2013.08.003