Research Article |
|
Corresponding author: Vellapandian Chitra ( dean.pharmacy.ktr@srmist.edu.in ) Corresponding author: Kaliappan Ilango ( ilangok67@gmail.com ) Academic editor: Ivan Dimitrov
© 2024 Anuragh Singh, Vellapandian Chitra, Kaliappan Ilango.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Singh A, Chitra V, Ilango K (2024) Network pharmacology exploration to reveal molecular insights of Phyllanthus niruri in non-alcoholic fatty liver: In vitro and in silico evidence. Pharmacia 71: 1-16. https://doi.org/10.3897/pharmacia.71.e124595
|
Abstract
The hepatic manifestation of metabolic syndrome, associated with various metabolic diseases such as type 2 diabetes, insulin resistance, and high cholesterol, is called non-alcoholic fatty liver disease (NAFLD). Despite several research efforts, no approved medicine is currently available for the treatment of this illness. Swiss Target Prediction was used to screen phytochemicals. To examine potential targets, the protein-protein interaction (PPI) network was developed. Cytoscape was used to create the component-target-pathway (C-T-P) network, and AutoDock was used to assess molecular docking. Antioxidant and anti-inflammatory qualities were tested in vitro. Naringenin, ellagic acid, and cyanidin were found to be the main active components. As important targets, PPARA, PPARG, and AKT1 were selected. Through enrichment analysis, a total of 20 crucial signaling pathways, including insulin resistance (IR), NAFLD, relaxin, PI3K-Akt, HIF-1, AGE-RAGE, and MAPK, were identified. The in silico computational techniques predicted the molecular pathway for the active ingredients and the disease targets, thus helping to further research.
Keywords
NAFLD, network pharmacology, gene ontology, KEGG, protein targets
Introduction
Non-alcoholic fatty liver disease (NAFLD) is the hepatic manifestation of metabolic syndrome, which is associated with a number of metabolic diseases, such as type 2 diabetes (T2DM), insulin resistance (IR), and hyperlipidemia (
Phyllanthus niruri is a coastal invasive plant belonging to the Euphorbiaceae family. It is also known as a windstorm or a stonebreaker. Its leaves and fruit have been used medicinally since antiquity (
The new field of network pharmacology examines disease mechanisms and medication action mechanisms in the context of larger biological networks (

Material and methods
Preparation of the extract and preliminary analysis
Prof. P. Jayaraman, taxonomists, plant anatomy, and research center authenticated the plant Phyllanthus niruri (PARC/2021/4526), and a voucher specimen was deposited in the department of pharmacognosy, SRM College of Pharmacy, for future reference. After authentication, the entire plant was cleaned systematically, shaded, and dried at room temperature. Dried leaves were separated from stalks and ground to a coarse powder using a porcelain mortar and pestle. Then the powder was weighed as 500 g dry powder, soaked in 70% ethanol and 30% distilled water, and subjected to Soxhlet for 72 hours at 60–70 °C. The filtrate was then concentrated and evaporated at 40 °C under reduced pressure using a rotary evaporator to obtain the crude extract. Phytochemical tests and other screenings were carried out on the extract of P. niruri to identify the constituents using standard procedures (
| Test | Standard | Results |
|---|---|---|
| Description | Brown colored fine powder | Pass |
| Loss on drying | NMT 7% w/w | 2.40% |
| Total Ash | NMT 20% w/w | 16.08% |
| Acid insoluble ash | NMT 7% | 1.20% |
| pH (1% w/v Aqueous) | 3–7 | 5.70 |
| Water soluble extractive | NLT 80% w/w | 89.35% |
| TLC | Complies | Complies |
| Total Bacterial content | NMT 10000 CFU/gm | 450 CFU/gm |
Determination of DPPH radical scavenging activity
Utilizing DPPH (2,2-diphenyl-1-picrylhydrazyl) free radicals, antioxidant activity in Phyllanthus niruri was calculated for free radical scavenging activity. One milliliter of DPPH solution (0.2 mM in methanol) was mixed with sample solutions at different concentrations (25, 50, 75, 100, and 200 µg/mL in methanol), and the reaction was allowed to carry out for 30 minutes at room temperature. After measuring the absorbencies of the solutions at 517 nm, the samples were examined for discoloration. Purple to yellow and pale pink were considered positive in strong and mild proportions, respectively (
To determine the sample’s IC50 value and the concentration of the sample required to inhibit 50% of the DPPH free radical, the log dosage inhibition curve was employed. The reaction mixture’s decreased absorbance revealed higher free radical activity. By contrasting the absorbance of each sample with that of a blank solution (a solution without any samples), the capacity of each sample to scavenge free radicals was calculated. Standard ascorbic acid was used as a point of comparison. The average findings (mean ± SD) of each analysis were obtained after it was done in triplicate.
The following equation calculated radical scavenging activity.
DPPH radical scavenging activity (%) = [(Absorbance of control - Absorbance of the test sample) / (Absorbance of control)] × 100.
Determination of the scavenging of superoxide radicals by alkaline DMSO
Scavenging of the superoxide (O2•-) anion radical was assessed by reducing nitroblue tetrazolium (NBT) using a previously published method (
% SO radical scavenging activity = (Control OD - Sample OD) × 100/Control OD.
Anti-inflammatory activity by inhibition of albumin denaturation
The reaction mixture comprises 2800 μL of phosphate-buffered saline, 200 μL of egg albumin or 450 μL of bovine serum albumin (5% w/v aqueous solution), and 1000 μL of plant extract (10–50 μg/mL). A small amount of 0.1N HCl was used to adjust the pH of the solution (6.3), which was heated to 57 °C for 30 minutes after the first 20-minute correction. The extracts in the suggested combination are replaced with distilled water as a negative control. After 15 minutes at 37 degrees, the mixtures are then incubated for 5 minutes at 70 degrees. Before measuring the solution’s absorbance at 660 nm, transfer the solution to a 96-well plate when it has been cooled. Diclofenac sodium was utilized as a standard (
The following formula calculates the percentage inhibition of albumin denaturation.
Percentage of inhibition denaturation (%) = [(A control – A sample) / A control] × 100
where A control is absorbance above all mixtures except drugs, and A sample is the absorbance reaction mixture with the sample.
Active substance screening and target prediction
The hydroalcoholic extract of Phyllanthus niruri (HAEPN) was subjected to gas chromatography-mass spectrometry (GC/MS) analysis using a Shimadzu 17A GC in conjunction with a Shimadzu QP2010 plus (quadrupole) Mass Spectrometer (Shimadzu, Japan), equipped with EI and a fused silica column DB-5 (30 m × 0.25 mm i.d.) of film thickness. The oven was preheated for 40 minutes at a temperature of 50–28,000 °C after being preheated for 5 minutes at 5000 °C. High-purity helium was used as a carrier gas in this experiment. The ionization voltage of the MS-analysis was adjusted to 70 eV using the EI method, the helium gas flow rate was set to 2 mL/min, and a split proportion of 1:30 mode was employed for sample injection of 1 μl. Using computer searches on the National Institute of Standards and Technology (NIST) Ver. 11 MS data library database and comparing the GC-MS spectrum to the spectrum of the known components stored in the NIST library, the names, molecular weights, and nature/structure of the phytoconstituents were confirmed (
Once the compound was selected through the GC/MS, the SMILES of the eluted compound were collected, and they were run on bulk analysis for ADMET screening through (SwissADME) (
NAFLD-related target screening
GeneCards (https://www.genecards.org) (
Potential target acquisition of HAEPN for NAFLD therapy
The genes obtained through Swiss TargetPrediction for the phytochemicals filtered from ADMET filtration were selected for interactions. Further, the genes obtained from the gene card and DisGiNET were co-related with those obtained from phytochemicals by drawing a Venn diagram (http://bioinformatics.psb.ugent.be/webtools/Venn/) and (https://bioinformatics.com.c) (
Establishing a protein-protein interaction network and identifying key targets
PPI analysis is critical for understanding complicated cell mechanisms and analyzing biological processes. The PPI network was built with STRING v11.5 (
Functional enrichment in gene ontology (GO) and pathway analysis in the Kyoto Encyclopaedia of Genes and Genomes (KEGG)
The Metascape platform (https://metascape.org/) was used to import putative HAEPN targets for NAFLD therapy (
Architecture of the compound-target pathway network
The compound-target and pathway network link for the treatment of non-alcoholic fatty liver disease (NAFLD) employing the HAEPN active ingredient, possible targets, and KEGG pathway information was established using the Cytoscape 3.7.2 software program. A set of topological measures, such as degree, betweenness, and closeness, were computed for the network. The Network Analyzer analytical method was used to identify the key target and substantial active components of HAEPN for the treatment of NAFLD.
Molecular docking
The key targets identified from HAEPN for NAFLD therapy and their matching active components were molecularly docked. The PubChem database (https://pubchem.ncbi.nlm.nih.gov/) was used to obtain compound structures, while the PDB database (https://www.rcsb.org/) downloaded core proteins. The proteins were screened through PharmaMapper (
The pedagogy of the concoction of macromolecules
The protein data bank (PDB) provided 3D protein structures in PDB format. The Biovia Discovery Studio Visualizer 2021 software was used to strip the proteins of ions, co-crystallized ligands, and water molecules. After that, missing hydrogens and Kollman partial charges were added to the grid in AutoDock. Tools 1.5.7 and non-polar hydrogens were combined with the matching carbons. A grid with the necessary size (60*60*60) and a grid spacing of 0.53 A° was created. The receptor was subsequently modified to be rigid and devoid of any flexible connections in order to undertake docking research. The structures were saved in PDB, partial charges (Q), and atom type (T) format files, also known as PDBQT protein receptor files, for use in future studies.
Preparation of the ligand file
The structure was created using ChemDraw Ultra 12.0 for a 2D structure after obtaining the ligand file from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/). The structure was then loaded into the Chem3D software for energy minimization using the MM2 force field. It was then translated into three-dimensional structures and saved as a PDB file. Then, ligands were made using the AutoDock tools 1.5.7 and M.G.L. tools 1.5.7. Hydrogen addition and protonation were used to alter the resulting 3D structures. To find the root, non-polar hydrogens were fused to the matching carbon rotations, and the program chose the torsion tree in the ligands as flexible. The structures were then stored as PDBQT ligand files for further study (
The construction of mating pockets
To construct mating spaces for docking in the grid design, the PDBQT receptor and ligand structures were opened in AutoDock Tools 1.5.7. When building a mating box, the sensitivity was set to 2.00, the spacing (angstrom) was set to 0.53 A°, the center was placed on the macromolecule, and the macromolecule’s properties in the x, y, and z dimensions in the center grid box were adjusted to maintain the protein covered by the mating pocket. These parameters include the coordinates of the mating box’s center (x, y, and z), its dimensions (x, y, and z), the maximum number of binding modes to be generated (set to 20), the energy range (set to 5), and the exhaustiveness (set to 8). To do more research, the output file was saved as a grid parameter file (GPF). A good level of 0 and the log results were used in the AutoGrid 4.2.6 run on the prepared GPF file. For a successful execution, the GLG file was examined.
Docking and visualization
The prepared macromolecule and ligand file were chosen, the search parameters were set to GA, the docking parameters were set to default, and the output was set to Lamarckian 4.2 and saved as a docking parameter file (DPF). The AutoDock was initiated with the DPF file at a nice level of 20. Upon completion of the docking process, the data files were saved in the formats pdbqt* and log*. To determine the binding affinity score between the target receptor proteins and small-molecule ligands, the docking binding free energy method was used using DLG data. In this study, docking models exhibiting the most minimal binding affinities were visualized, and the hydrogen bonds were represented in a two-dimensional format using Discovery Studio
Results
Phytochemical screening and preliminary analysis
The presence of alkaloids, carbohydrates, tannins, terpenoids, phenol, phlobatanins, steroids, cardiac glycosides, and reducing sugars was found in a hydroalcoholic extract of P. niruri; however, saponins were not found. The preliminary examination revealed that the loss on drying, total ash, acid insoluble ash, TLC, total bacterial count, pH, bulked, and tap density met Indian pharmacopoeia standards (Table
In vitro antioxidant study
Determination of DPPH radical scavenging activity
The absorbance at 517 nm measured using a UV-visible spectrophotometer was 0.06 for standard ascorbic acid and 0.04 for HAEPN, respectively. The IC50 values obtained were 10.89 and 10.27 µg/mL for ascorbic acid and HAEPN, respectively. That suggests that a higher concentration of hydroalcoholic extract from plants trapped more free radicals produced by DPPH, resulting in a drop in absorbance and an increase in percentage inhibition. The graph was plotted for the standard ascorbic acid (Fig.


Determination of the scavenging of superoxide radicals by alkaline DMSO
The ability to neutralize superoxide radicals in the reaction mixture using the test material at different dosages, compared to standard ascorbic acid, is shown by the decrease in absorbance at 560 nm. The IC50 values for ascorbic acid and HAEPN, which exhibit superoxide radical scavenging action, were determined to be 4.1715 μg/mL and 7.6499 μg/mL, respectively (Fig.

Anti-inflammatory activity by inhibition of albumin denaturation
HAEPN at concentrations ranging from 100 to 1000 µg/mL inhibited egg albumin denaturation, as evidenced by a concentration-dependent rise in solution absorbance. A comparable impact was seen with standard Diclofenac sodium concentrations ranging from 100 to 1000 µg/mL. HAEPN and diclofenac sodium had linear regression coefficients of R2 = 0.9906 and R2 = 0.9498, respectively, and the observed inhibition was dose dependent. The IC50 values of HAEPN and diclofenac sodium obtained from linear regression analysis were 64.30 µg/mL and 80.40 µg/mL, respectively. The plotted graph for the test sample is shown in Fig.


Active substance screening and target prediction
Fig.
| SI.No. | RT | Compound | Structure | MW | Formulae | Peak area % |
|---|---|---|---|---|---|---|
| 1 | 3.198 | Glycerin |

|
92.09 | C3H8O3 | 0.81 |
| 2 | 5.649 | 2-Furanmethanol |

|
98 | C5H6O2 | 0.43 |
| 3 | 7.100 | α-Angelica lactone |
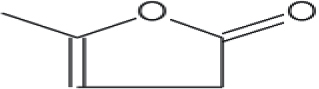
|
98 | C5H6O2 | 0.20 |
| 4 | 8.385 | 5,7-Dihydroxy-2-(4-hydroxyphenyl) chroman-4-one |

|
272 | C15H12O5 | 0.02 |
| 5 | 10.870 | (E)-1,2-ditert-butyldiazene |

|
142 | C8H18N2 | 0.34 |
| 6 | 11.267 | 4h-pyran-4-one, 3-hydroxy-2-methyl |
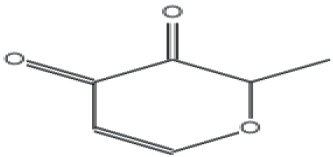
|
126 | C6H6O3 | 0.07 |
| 7 | 12.008 | 3,4-Dihydroxybenzoic acid |

|
154.1 | C7H6O4 | 0.09 |
| 8 | 20.337 | 1,2-Benzenedicarboxylic acid, diethyl ester |

|
222 | C12H14O4 | 6.13 |
| 9 | 27.416 | Ellagic acid |

|
302 | C14H6 O8 | 0.29 |
| 10 | 29.884 | 8,5’-Diferulic acid |
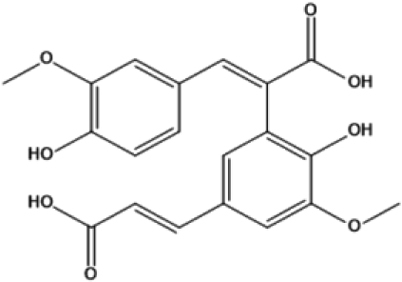
|
386 | C20H18O8 | 2.27 |
| 11 | 32.295 | 7-Hydroxy-6,9a-dimethyl-3-methylene-decahydro-azuleno[4,5-b] furan-2,9-dione |

|
264 | C15H20O4 | 1.06 |
| 12 | 33.867 | Quercetin-3-O-rhamnoside |

|
448 | C22H45Cl | 1.61 |
| 13 | 36.020 | Niruriflavone |
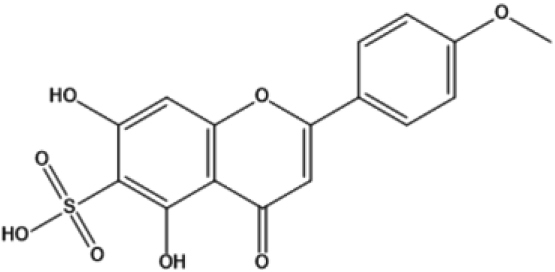
|
364 | C16H12O8S | 3.78 |
| 14 | 37.011 | 2-(3,4-Dihydroxyphenyl)-3,5,7-trihydroxy-1-Benzopyrylium |

|
282 | C15H11O6 + | 7.55 |
| 15 | 37.770 | 9-Bromononanoic acid |

|
237 | C9H17BrO2 | 0.80 |
| 16 | 38.927 | 18,19-Secoyohimban-19-oic acid, 16,17,20,21-tetradehydro-16-(hydroxymethyl)-, methyl ester |

|
352 | C21H24N2O3 | 1.93 |
| 17 | 39.220 | Isobornyl acrylate |

|
208 | C13H20O2 | 0.46 |

NAFLD-related target screening
A total of 1,808 and 1,058 genes were obtained through gene cards and DisGeNET for NAFLD-related targets with ‘Non-Alcoholic Fatty Liver Disorder’ and ‘NAFLD’ as keywords. The duplicates were removed, and 1,713 were selected for further evaluation. The overall unique elements were 2,482.
Potential targets of HAEPN for NAFLD therapy
There were 59 common targets found when a total of 1713 NAFLD-related targets intersected with 521 GCMS targets that included five main bioactive chemicals that were discovered during screening. For the GCMS, DisGeNET, and gene cards, a Venn diagram was drawn (Fig.

PPI network construction
The STRING database was used to import the common 59 targets for PPI network data. The PPI network has 374 edges and 59 nodes, with an average node degree of 12.7 and a local clustering coefficient of 0.576. The nodes in the network represent proteins, while the edges indicate PPI. The PPI enrichment value was found to be < 1.0e-16. The network obtained from STRING is shown in Fig.
The degree, average shortest path length, betweenness, and closeness centrality of the top 10 key targets.
| Target | Degree | Betweenness Centrality | Closeness Centrality | Avg Shortest Path Length |
|---|---|---|---|---|
| AKT1 | 41 | 0.111 | 0.777 | 1.285 |
| PPARG | 33 | 0.103 | 0.777 | 1.428 |
| VEGFA | 32 | 0.103 | 0.674 | 1.482 |
| EGFR | 30 | 0.045 | 0.658 | 1.517 |
| MMP9 | 29 | 0.039 | 0.658 | 1.517 |
| ESR1 | 28 | 0.033 | 0.643 | 1.553 |
| SIRT1 | 27 | 0.026 | 0.651 | 1.535 |
| PPARA | 26 | 0.038 | 0.629 | 1.589 |
| TLR4 | 26 | 0.037 | 0.602 | 1.660 |
| MMP2 | 22 | 0.007 | 0.589 | 1.696 |


GO functional enrichment and KEGG pathway analysis
The Metascape platform was used to conduct GO and KEGG pathway enrichment analyses on 59 putative HAEPN targets for NAFLD treatment. The obtained GO enrichment analysis results were of biological function, cellular function, and molecular function. The top ten GO characteristics were chosen, and the bioinformatics platform was used to record and visualize the data (Fig.

The bar diagram illustrates the GO enrichment analysis of genes associated with NAFLD. The study includes the top 20 enrichment keywords in three categories: biological processes (BP), cellular components (CC), and molecular functions (MF). The abscissa represents the proportion of genes of interest inside each entry, whereas the ordinate denotes the individual entries. There is a positive correlation between the size of a bar and the number of genes identified in the corresponding entry. The -LogP denotes the enrichment score associated with each ontology, as seen in the color bar.


Network construction analysis
The Compound-Target Pathway network diagram (Fig.

Molecular docking assessment
The best six proteins were selected for the analysis (3VI8, 4CI4, 3ET1, 5F9B, 1NFK, and 3TKM), and five selected active ingredients (i.e., naringenin, protocatechuic acid, diethyl phthalate, ellagic acid, and cyanidin) underwent molecular docking separately. After that, 30 sets of receptor-ligand docking outcomes were produced similarly. Intermolecular forces must be evaluated during the molecular docking process. In other words, the study’s intermolecular forces are mostly hydrogen bonds. Table
| Ligand | PPAR-A | PPAR-D | PPAR-G | ||
|---|---|---|---|---|---|
| 3VI8 | 4CI4 | 3ET1 | 3TKM | 5F9B | |
| Naringenin | -8.03 | -7.79 | -8.67 | -7.12 | -8.53 |
| Protocatechuic acid | -4 | -5.09 | -6.32 | -5.14 | -6.32 |
| Diethyl Phthalate | -5.4 | -5.27 | -5.34 | -6.05 | -6.87 |
| Cyanidin | -7.76 | -7.42 | -7.6 | -6.47 | -7.67 |
| Ellagic acid | -7.17 | -6.32 | -5.77 | -6.21 | -5.43 |
| Ligand | Target | Protein | Pocket Atom | 2D Interaction |
|---|---|---|---|---|
| Naringenin | PPAR α | 3ET1 |

|

|
| PPAR δ | 3TKM |

|

|
|
| PPAR γ | 5F9B |
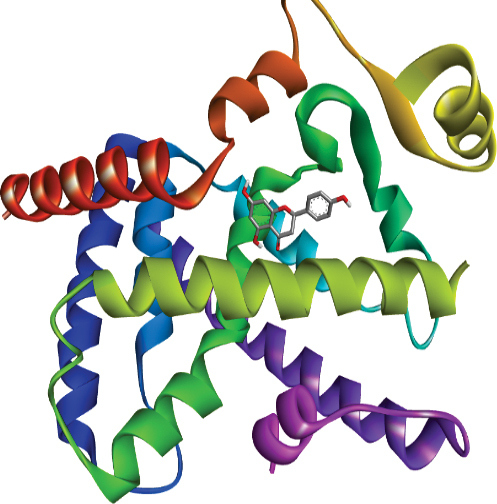
|

|






Discussion
Non-alcoholic steatohepatitis may develop over time as a silent state of non-alcoholic fatty liver disease. In the future, it may significantly increase the risk of cirrhosis, liver transplantation, and death. The major causes of central obesity and NAFLD include a sedentary lifestyle and excessive consumption of high-calorie foods such as butter, cheese, egg yolk, and lard fat (
These 17 phytoconstituents were subjected to SwissADME tool screening, and 5 compounds were screened based on Lipinski’s principle and lead likeness. The targets of putative compounds were integrated with the NAFLD-associated gene from the DisGeNET and gene card databases, and a common target plot was constructed. The common target was then used to form a PPI network, and KEGG and GO enrichment analyses were constructed. The network construction for the ligand and pathway with the gene was explored. The compound-target-pathway network was constructed (Fig.
According to certain studies, cyanidin and its glycosides are found in foods where lower levels of atherogenic lipoprotein and oxidized low-density lipoprotein (LDL) are produced both in vitro (
The study conducted by Arul and Subramanian in 2013 revealed that the growth of HepG2 cells was effectively suppressed by naringenin in a concentration-dependent manner. The activation of p53 has been associated with cell cycle arrest in both the G1 and G2 stages of the cell cycle. Significantly, the accumulation of p53 was enhanced by naringenin in a way that was dependent on the dosage. This observation provides a potential explanation for the G0/G1 and G2/M phase arrests induced by naringenin in HepG2 cells (
Notably, protocatechuic acid, ellagic acid, naringenin, cyanidin, and diethyl phthalate are the main active substances in HAEPN for the treatment of NAFLD. Such active ingredients may affect the PI3K-Akt, HIF-1, AGE-RAGE, and MAPK signaling pathways, as well as EGFR tyrosine kinase inhibitors, endocrine resistance, insulin resistance, non-alcoholic fatty liver disease, relaxin, and non-alcoholic fatty liver disease. The present research discusses a putative mechanism of action for HAEPN in the treatment of NAFLD utilizing network pharmacology and molecular docking technologies. This study also offers some conceptual and empirical support for the development of new NAFLD treatments.
Many constraints have plagued both molecular docking and network pharmacology. They can be utilized as a primary tool to sort out the active ingredient, specific targets, and their pathways through computational means. The currently accessible database has several uncertainties. At the same time, computation findings cannot completely replace experimental data; they must be combined with other methodologies and empirically confirmed in many circumstances for the logical design of polypharmacology (
To further corroborate our results, it is widely known that further studies such as hit/lead optimization, molecular dynamics, MMPBA (free energy calculation), and in vivo animal trials are required. Nevertheless, our breadth was constrained by time constraints in addition to other factors. Lead optimization comprises changing the chemical makeup of newly identified compounds to improve their toxicity, potency, and pharmacokinetics, thereby creating safer and more effective small molecules for the inhibition of NAFLD-relevant pathways.
Conclusion
Contrary to what was shown in the previous research, HAEPN did have higher levels of antioxidant and anti-inflammatory activity, thus creating a platform for further NAFLD activity augmentation. Nuclear receptor activity and ligand-induced transcription factor activity were shown to be significantly enriched in gene ontology’s molecular function. AKT1, PPARA, PPARG, MAPK8, GSK3B, INSR, NR1H3, and ERN1 all had improved connections in the PPI networks built for target proteins, suggesting they might be used to treat NAFLD. Based on our findings, we propose using network pharmacology, GO, KEGG, and molecular docking as the initial phase in doing this study. To develop a novel therapeutic agent for treating and managing NAFLD, we recommend further refining and enhancing the top two hits (naringenin and cyanidin). However, additional in vivo pharmacological research development of HAEPN and animal or cell line testing are needed to verify the reliability and rationality of the projected effects.
Acknowledgments
We thank the management, SRM College of Pharmacy, and SRMIST for providing facilities to carry out this work.
No funding was received to assist with the preparation of this manuscript.
References
- Abdel-Moemin AR (2011) Switching to black rice diets modulates low-density lipoprotein oxidation lipid measurements in rabbits. The American Journal of the Medical Sciences 341: 318–324. https://doi.org/10.1097/MAJ.0b013e3182019f62
- Abenavoli L, Milic N, Di Renzo L, Preveden T, Medić-Stojanoska M, De Lorenzo A (2016) Metabolic aspects of adult patients with nonalcoholic fatty liver disease. World Journal of Gastroenterology 22: 7006. https://doi.org/10.3748/wjg.v22.i31.7006
- Adedotun IO, Abdul-Hammed M, Hamzat BA, Adepoju AJ, Akinboade MW, Afolabi TI, Ismail UT (2022) Molecular docking, ADMET analysis, and bioactivity studies of phytochemicals from Phyllanthus niruri as potential inhibitors of hepatitis C virus NSB5 polymerase. Journal of the Indian Chemical Society 99: 100321. https://doi.org/10.1016/j.jics.2021.100321
- Al Zarzour RH, Ahmad M, Asmawi MZ, Kaur G, Saeed MaA, Al-Mansoub MA, Saghir SaM, Usman NS, Al-Dulaimi DW, Yam MF (2017) Phyllanthus niruri standardized extract alleviates the progression of non-alcoholic fatty liver disease and decreases atherosclerotic risk in Sprague–Dawley rats. Nutrients 9: 766. https://doi.org/10.3390/nu9070766
- Al Zarzour RH, Alshawsh MA, Asif M, Al-Mansoub MA, Mohamed Z, Ahmad M, Abdul Majid AMS, Asmawi MZ, Kaur G, Al-Dualimi DW (2018) Adipocytokine regulation and antiangiogenic activity underlie the molecular mechanisms of therapeutic effects of Phyllanthus niruri against non-alcoholic fatty liver disease. Nutrients 10: 1057. https://doi.org/10.3390/nu10081057
- Anuragh S, Ilango K (2022) Role of medicinal plants and their chemical constituents in ameliorating the cause for non-alcoholic fatty liver disorder—A Review. Journal of Applied Pharmaceutical Science 12: 043–055. https://doi.org/10.7324/JAPS.2022.120605
- Arul D, Subramanian P (2013) Naringenin (Citrus flavonone) induces growth inhibition, cell cycle arrest and apoptosis in human hepatocellular carcinoma cells. Pathology & Oncology Research 19: 763–770. https://doi.org/10.1007/s12253-013-9641-1
- Asadujjaman M, Hossain MA, Karmakar UK (2013) Assessment of DPPH free radical scavenging activity of some medicinal plants. Pharmacology Online 1: 161–165.
- Bardou P, Mariette J, Escudié F, Djemiel C, Klopp C (2014) jvenn: an interactive Venn diagram viewer. BMC Bioinformatics 15: 1–7. https://doi.org/10.1186/1471-2105-15-293
- Bessone F, Razori MV, Roma MG (2019) Molecular pathways of nonalcoholic fatty liver disease development and progression. Cellular and Molecular Life Sciences 76: 99–128. https://doi.org/10.1007/s00018-018-2947-0
- Colpo E, Vilanova CD, Pereira RP, Reetz LGB, Oliveira L, Farias IL, Boligon AA, Athayde ML, Rocha JBT (2014) Antioxidant effects of Phyllanthus niruri tea on healthy subjects. Asian Pacific Journal of Tropical Medicine 7: 113–118. https://doi.org/10.1016/S1995-7645(14)60005-5
- Consortium U (2019) UniProt: a worldwide hub of protein knowledge. Nucleic Acids Research 47: D506–D515. https://doi.org/10.1093/nar/gky1049
- Daina A, Michielin O, Zoete V (2017) SwissADME: a free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Scientific Reports 7: 42717. https://doi.org/10.1038/srep42717
- Dallas C, Gerbi A, Tenca G, Juchaux F, Bernard F-X (2008) Lipolytic effect of a polyphenolic citrus dry extract of red orange, grapefruit, orange (SINETROL) in human body fat adipocytes. Mechanism of action by inhibition of cAMP-phosphodiesterase (PDE). Phytomedicine 15: 783–792. https://doi.org/10.1016/j.phymed.2008.05.006
- Décordé K, Teissèdre PL, Sutra T, Ventura E, Cristol JP, Rouanet JM (2009) Chardonnay grape seed procyanidin extract supplementation prevents high‐fat diet‐induced obesity in hamsters by improving adipokine imbalance and oxidative stress markers. Molecular Nutrition & Food Research 53: 659–666. https://doi.org/10.1002/mnfr.200800165
- Dhanaraj CJ, Jebapriya M (2021) Synthesis, structural elucidation, biological activities of biomaterial containing biologically active ligand derived from Schiff base. International Journal of Mechanical Engineering 6(3): 3072–3080.
- Etl F, Kaiser C, Reiser O, Schubert M, Dötterl S, Schönenberger J (2022) Evidence for the recruitment of florivorous plant bugs as pollinators. Current Biology 32: 4688–4698. https://doi.org/10.1016/j.cub.2022.09.013
- Fontana M, Mosca L, Rosei MA (2001) Interaction of enkephalins with oxyradicals. Biochemical Pharmacology 61: 1253–1257. https://doi.org/10.1016/S0006-2952(01)00565-2
- Gfeller D, Grosdidier A, Wirth M, Daina A, Michielin O, Zoete V (2014) SwissTargetPrediction: a web server for target prediction of bioactive small molecules. Nucleic Acids Research 42: W32–W38. https://doi.org/10.1093/nar/gku293
- Harishchander A (2017) A computational approach for identifying the impact of pharmacogenomics in regulatory networks to understand disease pathology. Journal of Bioengineering and Biomedical Sciences 7: 2. https://doi.org/10.4172/2155-9538.1000235
- Hernández-Aquino E, Quezada-Ramírez MA, Silva-Olivares A, Casas-Grajales S, Ramos-Tovar E, Flores-Beltrán RE, Segovia J, Shibayama M, Muriel P (2019) Naringenin attenuates the progression of liver fibrosis via inactivation of hepatic stellate cells and profibrogenic pathways. European Journal of Pharmacology 865: 172730. https://doi.org/10.1016/j.ejphar.2019.172730
- Hsin K-Y, Ghosh S, Kitano H (2013) Combining machine learning systems and multiple docking simulation packages to improve docking prediction reliability for network pharmacology. PLOS ONE 8: e83922. https://doi.org/10.1371/journal.pone.0083922
- Jia Y, Kim J-Y, Jun H-J, Kim S-J, Lee J-H, Hoang MH, Kim HS, Chang HI, Hwang K-Y, Um S-J (2013) Cyanidin is an agonistic ligand for peroxisome proliferator-activated receptor-alpha reducing hepatic lipid. Biochimica et Biophysica Acta (BBA)-Molecular and Cell Biology of Lipids 1831: 698–708. https://doi.org/10.1016/j.bbalip.2012.11.012
- Jiao X, Jin X, Ma Y, Yang Y, Li J, Liang L, Liu R, Li Z (2021) A comprehensive application: Molecular docking and network pharmacology for the prediction of bioactive constituents and elucidation of mechanisms of action in component-based Chinese medicine. Computational Biology and Chemistry 90: 107402. https://doi.org/10.1016/j.compbiolchem.2020.107402
- Meyer AS, Heinonen M, Frankel EN (1998) Antioxidant interactions of catechin, cyanidin, caffeic acid, quercetin, and ellagic acid on human LDL oxidation. Food Chemistry 61: 71–75. https://doi.org/10.1016/S0308-8146(97)00100-3
- Milić S, Štimac D (2012) Nonalcoholic fatty liver disease/steatohepatitis: epidemiology, pathogenesis, clinical presentation and treatment. Digestive Diseases 30: 158–162. https://doi.org/10.1159/000336669
- Niu M, Zhang S, Zhang B, Yang K, Li Z (2021) Interpretation of Guideline for Web-Based Pharmacologic Evaluation methods. Chin. Trad. Herb. Drugs. LI Shao (Interpretation of Network Pharmacology Evaluation Method Guidance) 52: 4119–4129.
- Otasek D, Morris JH, Bouças J, Pico AR, Demchak B (2019) Cytoscape automation: empowering workflow-based network analysis. Genome Biology 20: 1–15. https://doi.org/10.1186/s13059-019-1758-4
- Rahman F, Tabrez S, Ali R, Alqahtani AS, Ahmed MZ, Rub A (2021) Molecular docking analysis of rutin reveals possible inhibition of SARS-CoV-2 vital proteins. Journal of Traditional and Complementary Medicine 11: 173–179. https://doi.org/10.1016/j.jtcme.2021.01.006
- Sakthivel B, Nisha SA (2019) Assessment of Nutritional composition antioxidant activity of edible herbs Solanum nigrum and Phyllanthus niruri. International Journal of Research Instinct 6(1): 1–12.
- Shaikh JR, Patil M (2020) Qualitative tests for preliminary phytochemical screening: An overview. International Journal of Chemical Studies 8: 603–608. https://doi.org/10.22271/chemi.2020.v8.i2i.8834
- Shiri-Shahsavar MR, Alijani S, Parsamanesh N, Nazari SS, Moazzen S, Majnouni A, Sadeghi N, Rasouli A (2023) The effect of grape-seed oil on diabetes-related hyperglycemia, dyslipidemia, and inflammation in streptozotocin-induced diabetic rats. Obesity Medicine 37: 100476. https://doi.org/10.1016/j.obmed.2022.100476
- Singh A, Dhanasekaran D, Ganamurali N, Preethi L, Sabarathinam S (2021) Junk food-induced obesity-a growing threat to youngsters during the pandemic. Obesity Medicine 26: 100364. https://doi.org/10.1016/j.obmed.2021.100364
- Siva Kumar B, Anuragh S, Kammala AK, Ilango K (2022) Computer aided drug design approach to screen Phytoconstituents of Adhatoda vasica as potential inhibitors of SARS-CoV-2 Main protease enzyme. Life 12: 315. https://doi.org/10.3390/life12020315
- Sivell C (2019) Nonalcoholic fatty liver disease: a silent epidemic. Gastroenterology Nursing 42: 428–434. https://doi.org/10.1097/SGA.0000000000000443
- Soni A, Sosa S (2013) Phytochemical analysis and free radical scavenging potential of herbal and medicinal plant extracts. Journal of Pharmacognosy and Phytochemistry 2: 22–29.
- Stelzer G, Rosen N, Plaschkes I, Zimmerman S, Twik M, Fishilevich S, Stein TI, Nudel R, Lieder I, Mazor Y (2016) The GeneCards suite: from gene data mining to disease genome sequence analyses. Current Potocols in Bioinformatics 54: 1.30.31–31.30.33. https://doi.org/10.1002/cpbi.5
- Szklarczyk D, Gable AL, Lyon D, Junge A, Wyder S, Huerta-Cepas J, Simonovic M, Doncheva NT, Morris JH, Bork P (2019) STRING v11: protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Research 47: D607–D613. https://doi.org/10.1093/nar/gky1131
- Tona MR, Tareq AM, Sayeed MA, Mahmud MH, Jahan I, Sakib SA, Shima M, Emran TB (2020) Phytochemical screening and in vitro pharmacological activities of methanolic leaves extract of Caryota mitis. Journal of Advanced Biotechnology and Experimental Therapeutics 3: 109–115. https://doi.org/10.5455/jabet.2020.d114
- Ugbaja RN, Akinloye DI, James AS, Ugwor EI, Kareem SE, David G, Abolade SO, Oyebade OE (2020) Crab derived dietary chitosan mollifies hyperlipidemia-induced oxidative stress and histopathological derangements in male albino rats. Obesity Medicine 20: 100300. https://doi.org/10.1016/j.obmed.2020.100300
- Wang X, Shen Y, Wang S, Li S, Zhang W, Liu X, Lai L, Pei J, Li H (2017) PharmMapper 2017 update: a web server for potential drug target identification with a comprehensive target pharmacophore database. Nucleic Acids Research 45: W356–W360. https://doi.org/10.1093/nar/gkx374
- Wu F, Liu J, Cao Z, Wang T, Ye L, Zhu M, Wang Z (2022) Molecular Mechanism of the Saposhnikovia divaricata–Angelica dahurica Herb Pair in Migraine Therapy Based on Network Pharmacology and Molecular Docking. Evidence-Based Complementary and Alternative Medicine 2022. https://doi.org/10.1155/2022/1994575
- Yao Y, Pan C, Wang C, Jing X (2020) Research Progress and application strategies network pharmacology in Traditional Chinese Medicine compounds. Journal of Chengdu University (Natural Science Edition) 39: 257–263.
- Zhou Y, Zhou B, Pache L, Chang M, Khodabakhshi AH, Tanaseichuk O, Benner C, Chanda SK (2019) Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nature Communications 10: 1523. https://doi.org/10.1038/s41467-019-09234-6
- Zhu Y-W, Yan X-F, Ye T-J, Hu J, Wang X-L, Qiu F-J, Liu C-H, Hu X-D (2021) Analyzing the potential therapeutic mechanism of Huashi Baidu Decoction on severe COVID-19 through integrating network pharmacological methods. Journal of Traditional and Complementary Medicine 11: 180–187. https://doi.org/10.1016/j.jtcme.2021.01.004